
Echinopericlimenes hertwigi ( Balss, 1913 ) Balss, 1913
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3835.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:A2720275-3774-4D4D-9D85-1E088C817619 |
|
DOI |
https://doi.org/10.5281/zenodo.6136323 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC1526-AE5A-FF96-FF76-FD8BFAFBDC6E |
|
treatment provided by |
Plazi |
|
scientific name |
Echinopericlimenes hertwigi ( Balss, 1913 ) |
| status |
comb. nov. |
Echinopericlimenes hertwigi ( Balss, 1913) View in CoL comb. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 15 View FIGURE 15 )
Periclimenes hertwigi Balss, 1913: 235 View in CoL [type locality: Sagami Nada, Japan, 120 m].— Balss, 1914b:49, figs. 28-30. Periclimenes (Ancylocaris) gracilirostris Kubo, 1940: 41 View in CoL , figs. 8–10 [type locality: Kumano Nada off Mie Prefecture, Japan, about 310 m].
Material examined. Japan: 1 ovigerous female (pcl 4.8 mm) (CBM-ZC-3692), Sakushita, Ohse-zaki, Suruga Bay, depth 45 meters, associated with sea urchin, SCUBA diving, 11 Apr. 1996. New Caledonia: 1 female (with bopyrid) (pcl. 4.8 mm) (MNHN-Na11143), SMIB 2, stn. DW6, 22°56’S, 167°16’E, depth 442–460 meters, 17 Sept. 1986; 1 male (pcl. 4.0 mm) (MNHN-Na11156), CHALCAL 2, stn. DW75, 24°39.31’S, 168°39.67’E, depth 600 meters, 29 Oct. 1986; 1 male (pcl. 6.9 mm), 1 ovigerous female (pcl. 7.3 mm) (MNHN-Na12050), CARLSUB, dive 2, 22°45S, 167°09E, Isle de Pines, depth 344–330 meters, 12 Mar. 1989.
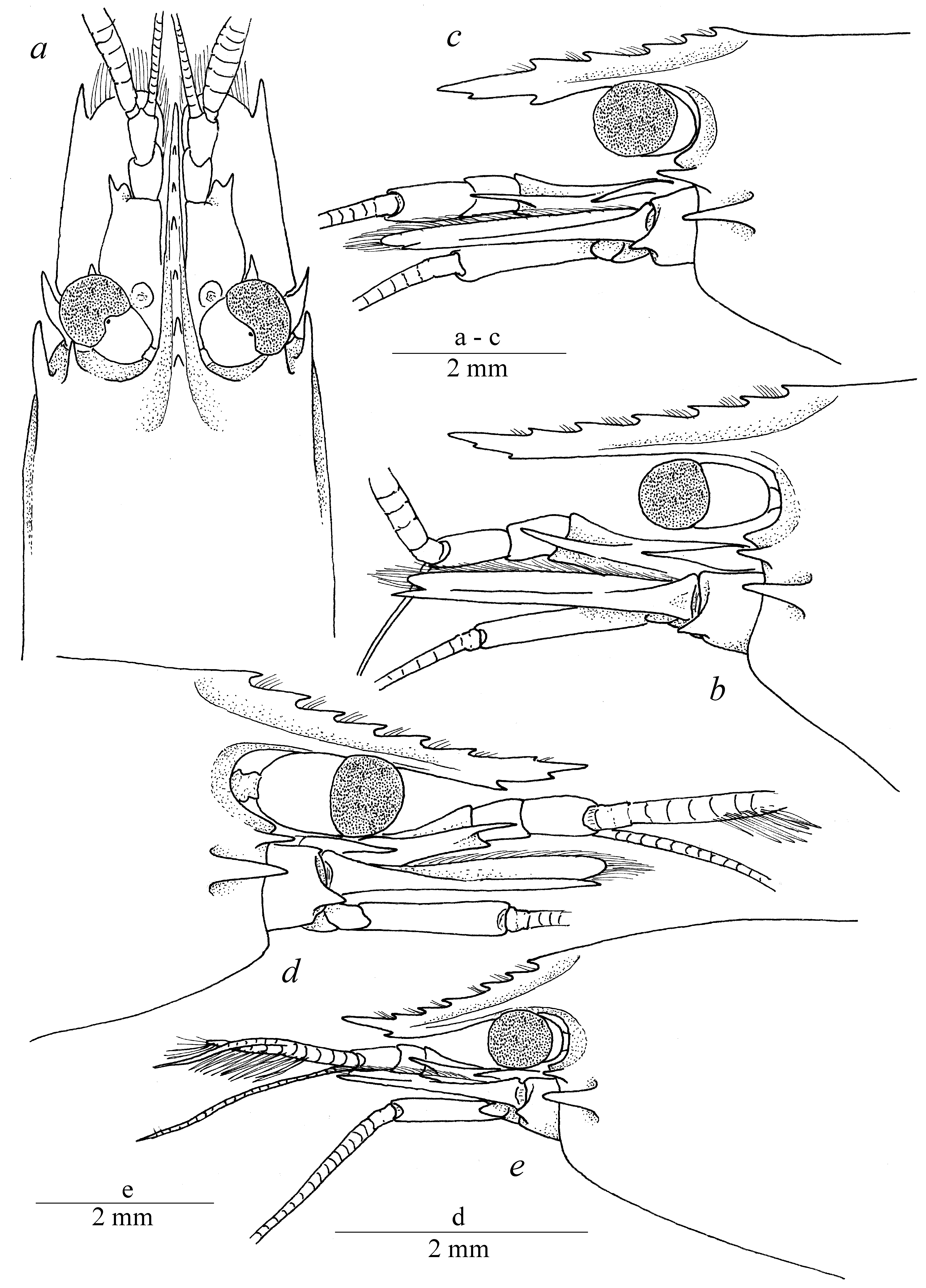
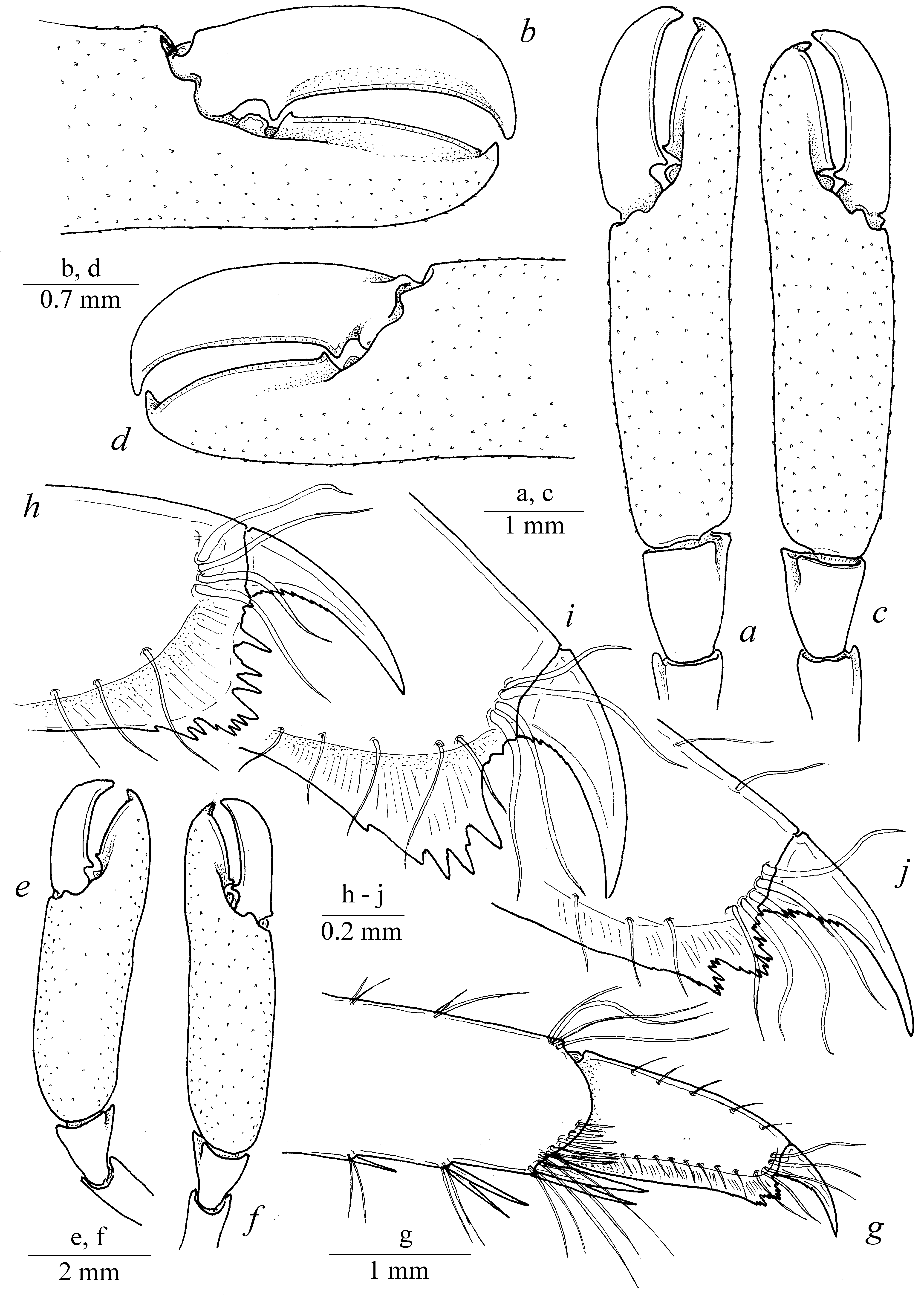
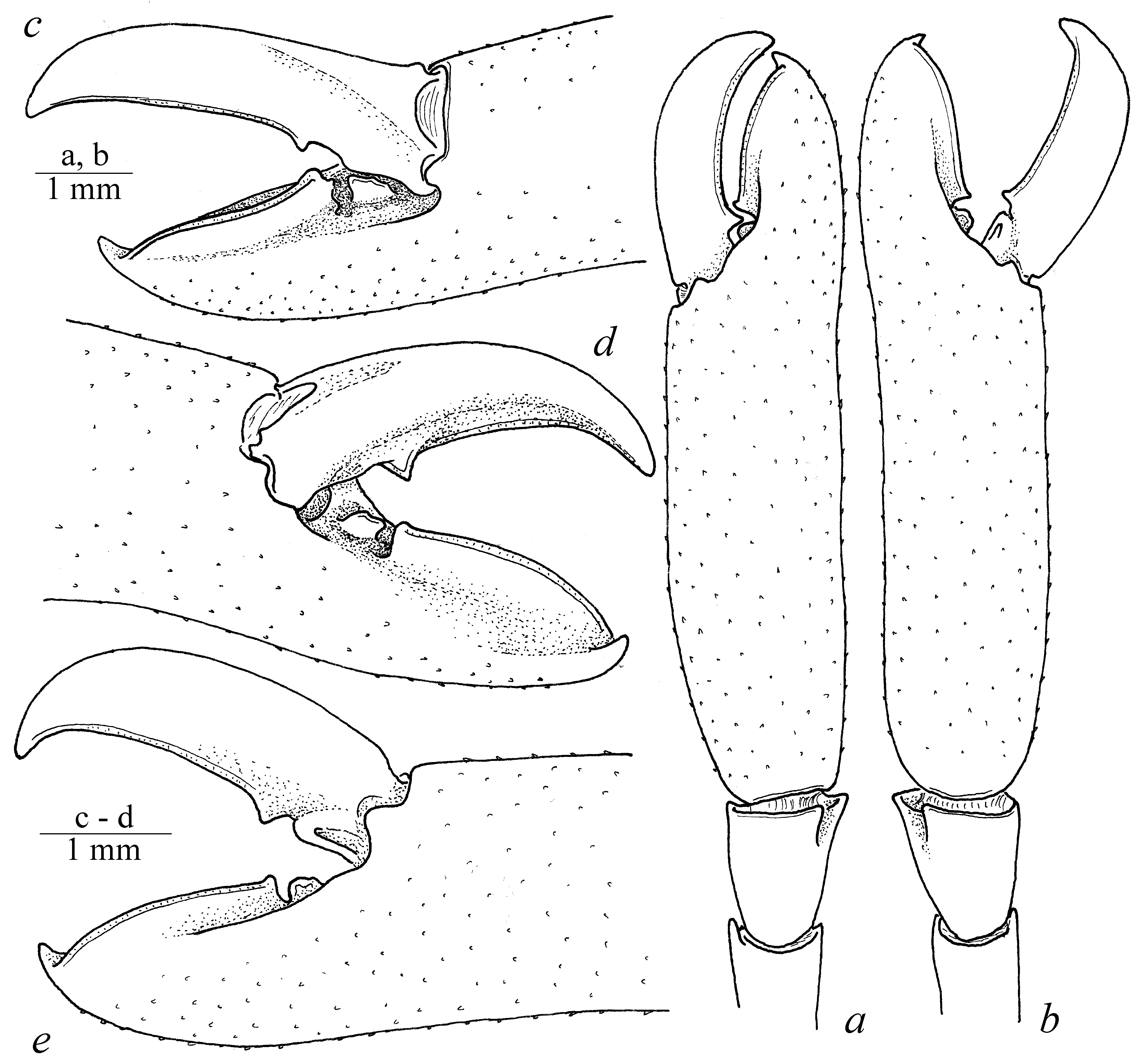
Diagnosis. Large-sized shrimps with subcylindrical body ( Fig. 1 View FIGURE 1 ). Carapace smooth, not pitted, swollen, with antennal and hepatic teeth; hepatic tooth large, situated close to pterygostomial margin of carapace ( Fig. 2 View FIGURE 2 ). Rostrum long and slender, compressed, directed downward, overreaching antennular peduncle; dorsal carina with 7–8 teeth, most proximal tooth situated slightly behind level of orbit, distal part unarmed; ventral margin feebly developed, with 2 small teeth situated at distal third, proximal ventral tooth situated at level of most distal dorsal tooth; tip of rostrum sharply produced; proximal lateral lamina feebly developed ( Fig. 2 View FIGURE 2 a); pterygostomial angle rounded. Abdominal somites smooth; pleurae of abdominal somites I–V rounded ( Fig. 1 View FIGURE 1 ). Telson stout, about twice longer than proximal width, narrowing distally, with 2 pairs of small dorsal marginal spines at 0.5 and 0.7 of telson length; distal margin armed with 3 pairs of spines including 1 pair of short stout lateral spines, 1 pair of long slender intermediate spines and 1 pair of simple medial spines about half intermediate spine length. Eyes ( Fig. 2 View FIGURE 2 ) large, with large hemispherical cornea and stout conical eyestalk. Antennula well developed, compressed; basal segment with distolateral angle bearing large tooth and medial rounded projection, with small ventromedial tooth. Antenna normal, basicerite armed with large lateral tooth overreaching distal margin of segment, with long slender carpocerite; scaphocerite wide, armed with sharp distolateral tooth slightly overreaching blade. Mandible robust, without palp; with well developed incisor process bearing 3–4 terminal teeth; molar process robust, with stout sharp distal teeth. Maxillula normal, with well developed palp bearing large triangular pointed dorsal lobe; upper lacinia wide, slightly curved, flaring distally, with strong setae along distal margin; lower lacinia more slender, tapering distally, covered with simple setae. Maxilliped III robust, with well developed exopod slightly overreaching antepenultimate segment; penultimate segment about 4 times as long as wide; distal segment triangular, with stout simple setae; epipod rounded, curved; small arthrobranch present. Pereiopod I slender; palm about twice longer than wide, as long as fingers; dactylus and fixed finger (pollex) simple, with entire cutting margins and simple tips; pollex furnished with several tufts of simple setae along ventral margin. Pereiopods II similar in shape, equal in size ( Figs. 2 View FIGURE 2 a, c, e, f; 4 a, b); merus and ischium slender, armed with small teeth along ventral margin, smooth dorsally; carpus small, triangular in shape, slightly overlapping carpo-propodal articulation, with straight margins; propodus cylindrical, covered with small blunt tubercles, about 3 times as long as wide; fixed finger (pollex) about twice as long as wide, covered with blunt tubercles along ventral margin, with large triangular tooth proximally, cutting edge slightly convex distally, with simple curved tip; movable finger (dactylus) relatively stout, slightly longer than polex, with large triangular tooth proximally, cutting edge slightly concave distally, tip simple and curved ( Figs. 3 View FIGURE 3 b, d; 4 c–e). Pereiopod III relatively slender, with unarmed proximal segments; carpus slender, about 4 times as long as wide, unarmed, distodorsal margin of the segment bluntly produced and overlapping carpo-propodal articulation; propodus slender, about 7 times as long as wide, with straight lateral margins, with 5 slender spines along ventral margin and several slender spines at distoventral angle ( Fig. 3 View FIGURE 3 g), with tuft of stout flattened setae at dactylo-propodal articulation; dactylus ( Fig. 3 View FIGURE 3 g) relatively robust, about twice longer than wide, ventral margin straight bearing several strong teeth distally; unguis small and curved, proxy-ventrally serrated, with distal part unarmed ( Fig. 3 View FIGURE 3 h–j). Uropods normal, well developed; distolateral angle of exopod sharply produced, with slender movable spine.
Remarks. The present specimens are identical to the original description of the species given by Balss (1914) and re-description presented by Kubo (1940). Echinopericlimenes hertwigi comb. nov. can be readily separated from the congeners by having relatively long rostrum usually armed with 2 ventral teeth, with the proximal ventral tooth situated around the level of the most distal dorsal rostral tooth. Some differences in the shape of rostrum can be found between the examined specimens from Japan and New Caledonia. Rostrum of female from Japan ( Fig. 2 View FIGURE 2 e) is strongly sloping downward, concave at its medial part, with 1 ventral tooth situated at the level of the most distal dorsal tooth. Nevertheless, rostrum is nearly straight, slightly sloping downward, with 2 ventral teeth in the specimens from New Caledonia ( Fig. 2 View FIGURE 2 a–d). Moreover, Japanese specimen was collected from the depth of 45–310 meters (see Balss, 1913; Kubo, 1940) while New Caledonian specimens were collected deeper, from 330–600 meters; specimens were collected from different hosts (see below). An attempt to compare the sequences of barcoding gene COI between these specimens failed due to the inappropriate preservation. Newly collected specimens from these regions eligible for genetic sequencing may prove if the specimens from such distant areas conspecific or belong to separate species.
Coloration. Color information is available for this species from some photos taken in shallow waters off Honshu Island posted on the Internet (see Fig. 15 View FIGURE 15 ) and the labeled description of alive coloration of the ovigerous female from Suruga Bay in the present study (CBM-ZC-3692): “ body and proximal segments of appendages uniformly red; dorsal surface of fingers, dorsomesial part of the palm, dorsal surface of carpus and distodorsal margin of merus of pereiopod II with white pigmentation; distal margin of meri, carpi, propodi and dactyli of ambulatory pereiopods white; white longitudinal band covering dorsal margin of abdominal somites II–III extending to the proximal part of telson ”.
Hosts. Host records were presented by Balss (1913, 1914) and Bruce (1983; 1991) and underwater photographs on Japanese specimens ( Fig. 15 View FIGURE 15 ). The holotype was found in association with deep-water sea urchin of the genus Phormosoma Rhomson, 1872 (Echinothuriidae) (after Balss, 1913, 1914), while a pair of specimens from New Caledonia (MNHN-Na12050) were associated with deep-water sea urchin of the genus Asthenosoma Grube, 1868 (Echinothuriidae) ( Bruce, 1991). Bruce (1983) also recorded the species from Queensland, Australia as a symbiont of Araeosoma thetidis (H.L. Clark, 1909) (Echinothuriidae) . In relatively shallow waters near Honsu Island, Japan, the species is often found in association with deep-water venomous sea urchin A. thetidis ( Fig. 15 View FIGURE 15 ) and Araeosoma owstoni Mortensen, 1904 (see Debelius, 1999). Possibly host sea urchins migrate to shallow waters following the cold upwelling currents as all records of E. hertwigi at shallow waters off Japan were done during the winter season.
Distribution. The species is known off Honsu Island, Japan, 45–310 meters (the type locality; Balss, 1913; Kubo, 1940); Indonesia, 304 meters (Holthuis, 1952); Queensland, Australia, 275 meters ( Bruce, 1983) and New Caledonia, 334–600 meters ( Bruce, 1990, 1991). Nevertheless, distribution of E. hertwigi comb. nov. needs to be verified as not all known specimens of the species were re-examined during the present study as well as some minor morphological differences are reported for the specimens from distant localities (see Remarks above).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
SubFamily |
Pontoniinae |
|
Genus |
Echinopericlimenes hertwigi ( Balss, 1913 )
| Marin, Ivan & Chan, Tin-Yam 2014 |
Periclimenes hertwigi
| Kubo 1940: 41 |
| Balss 1914: 49 |
| Balss 1913: 235 |