
Paramicromerys, MILLOT, 1946
|
publication ID |
https://doi.org/10.1046/j.1096-3642.2003.00046.x |
|
persistent identifier |
https://treatment.plazi.org/id/03F987F6-FFEE-FFA4-FC64-0BAF3001F9DC |
|
treatment provided by |
Carolina |
|
scientific name |
Paramicromerys |
| status |
|
PARAMICROMERYS MILLOT, 1946 View in CoL View at ENA
Paramicromerys Millot, 1946: 145 View in CoL ; type species by original designation: Micromerys madagascariensis Simon, 1893 .
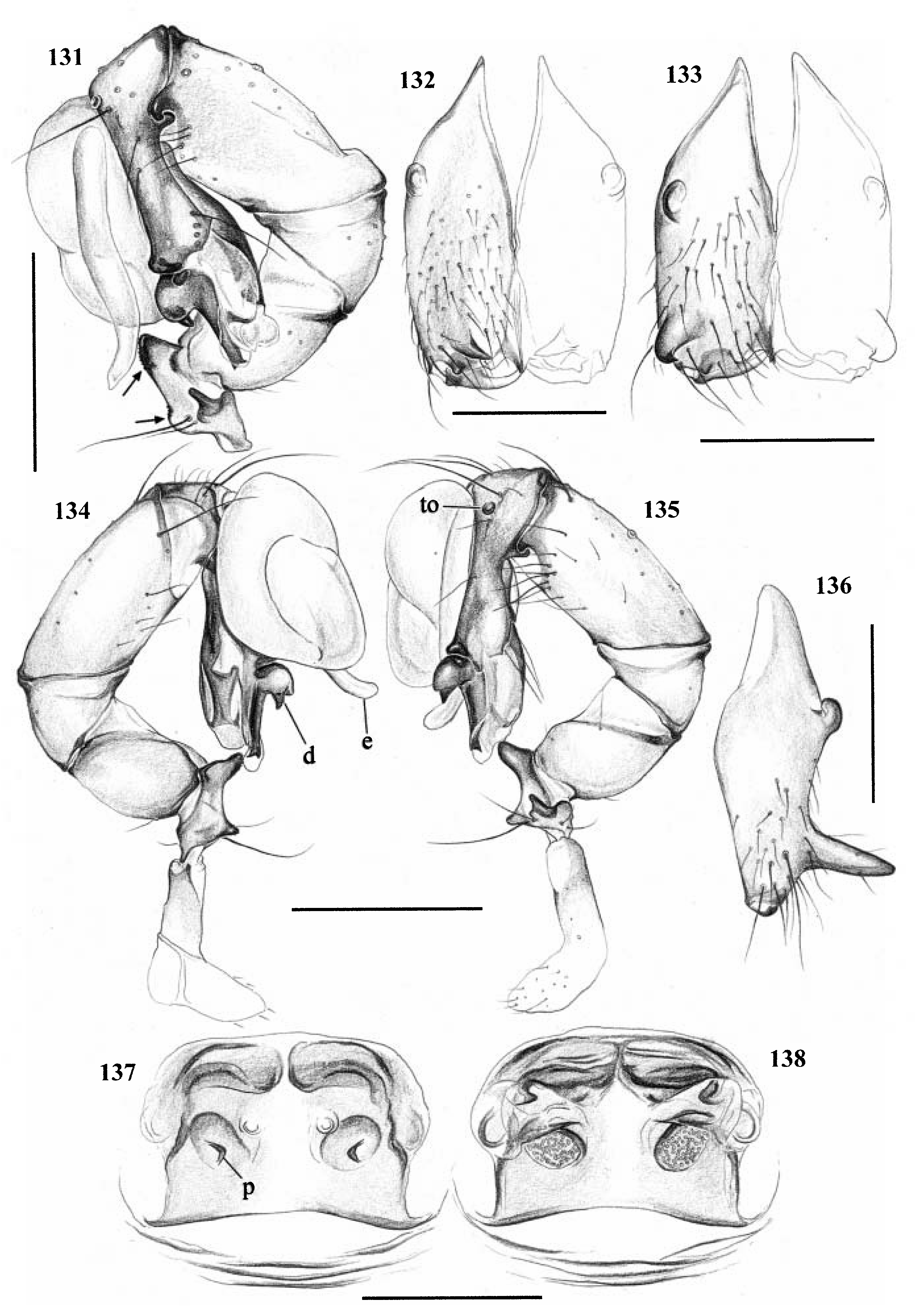
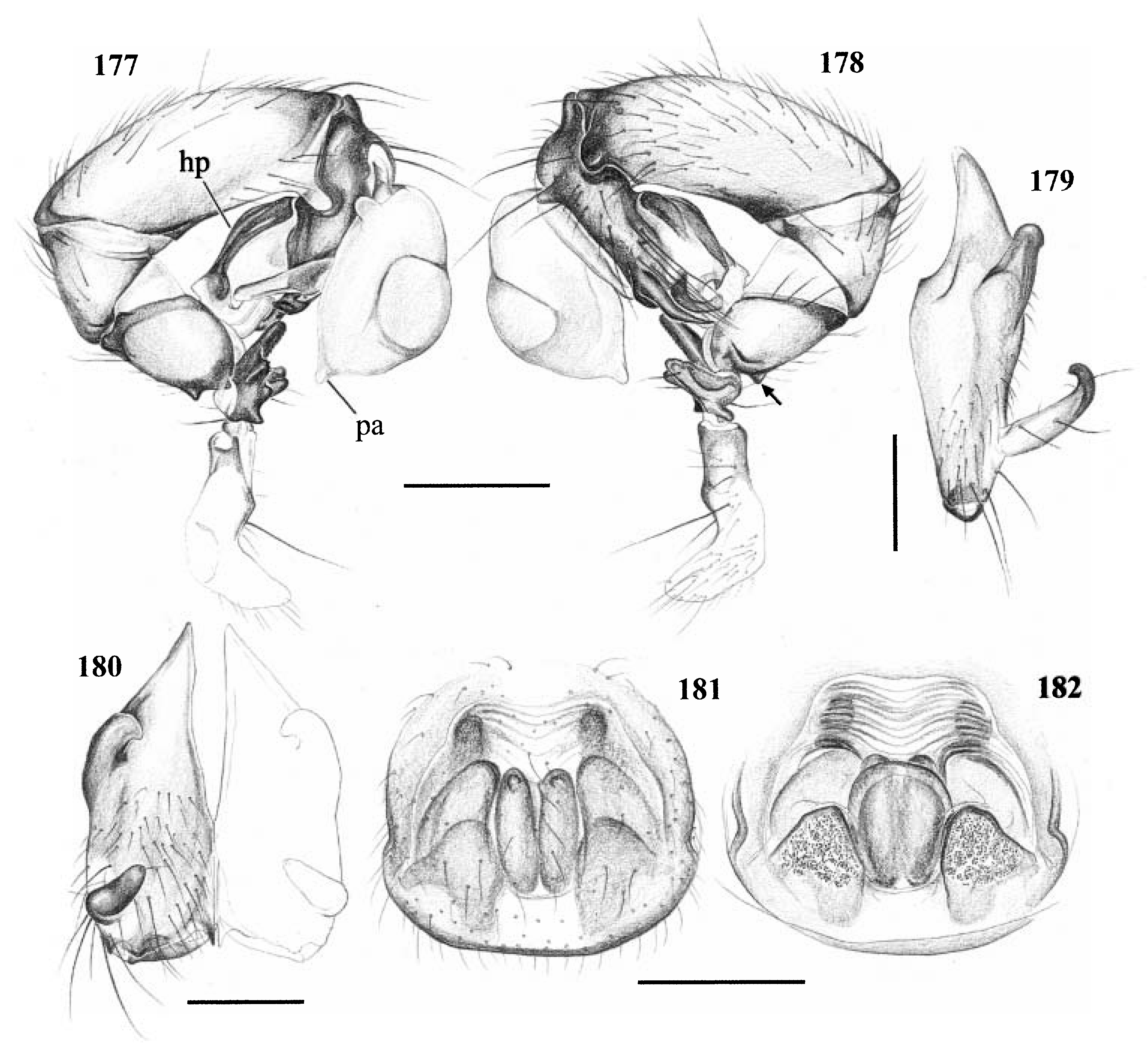
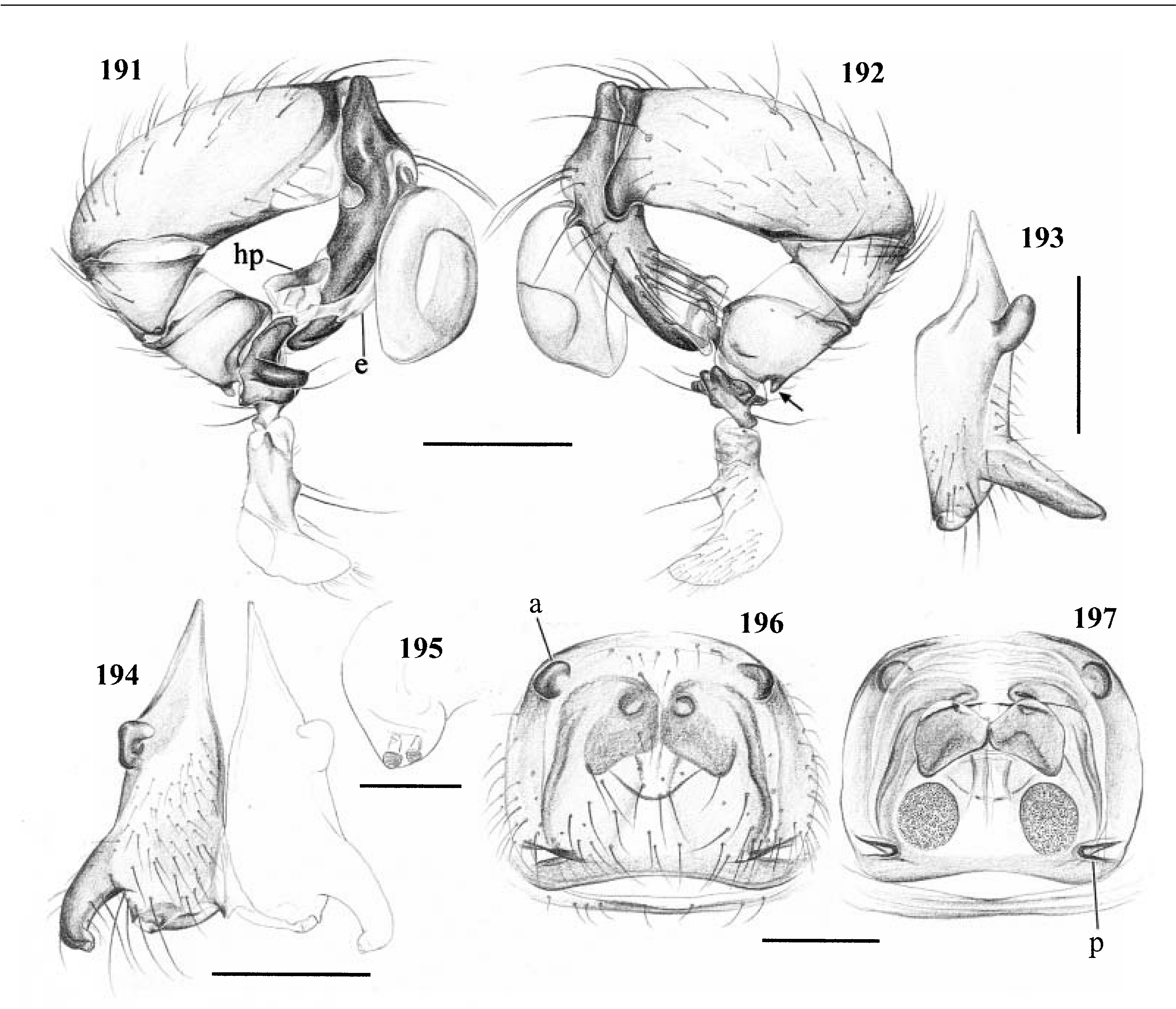
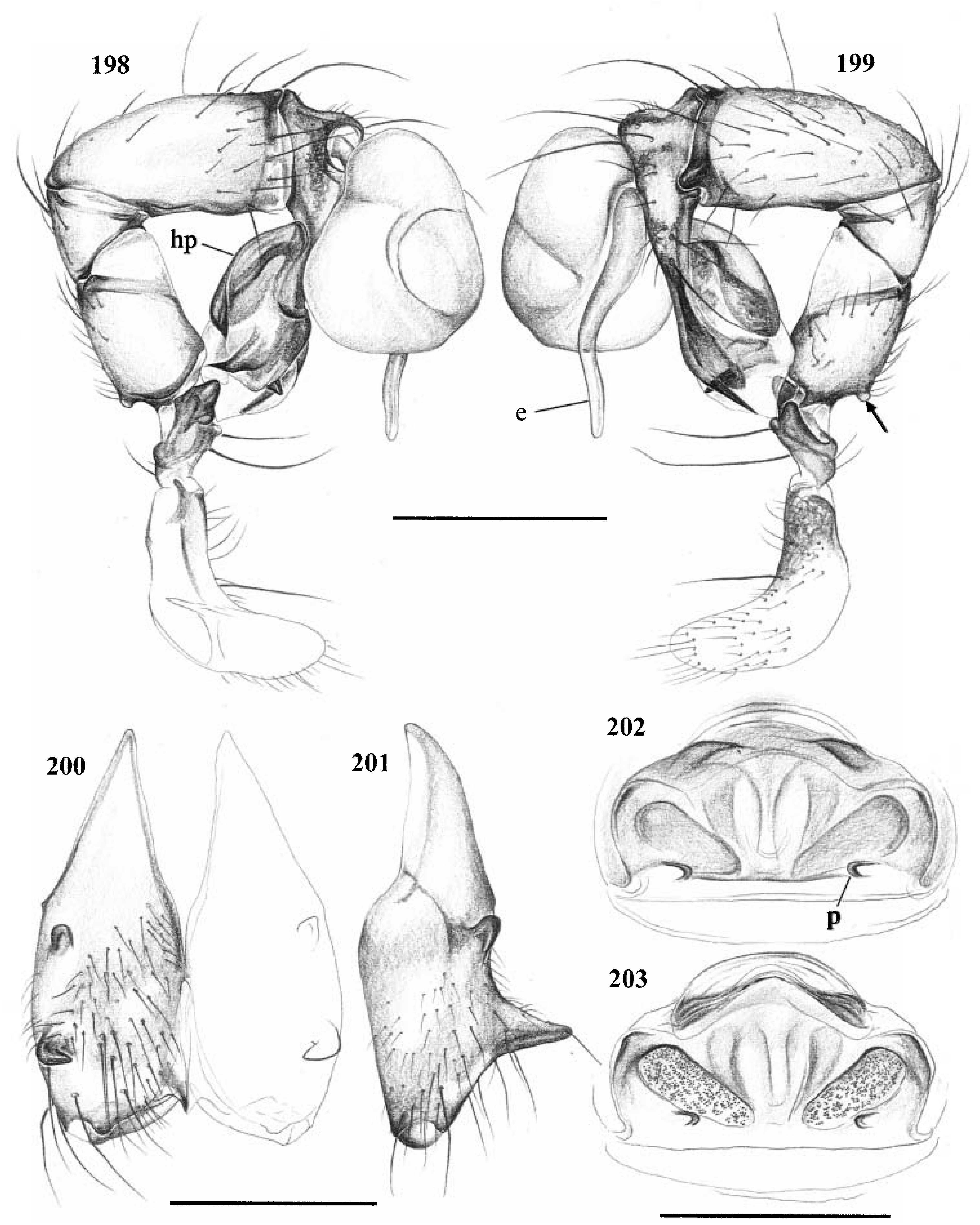
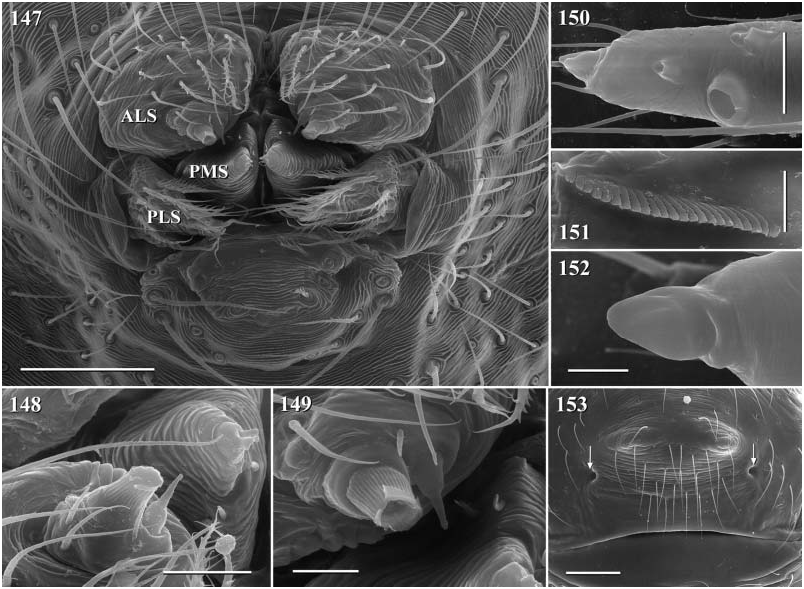
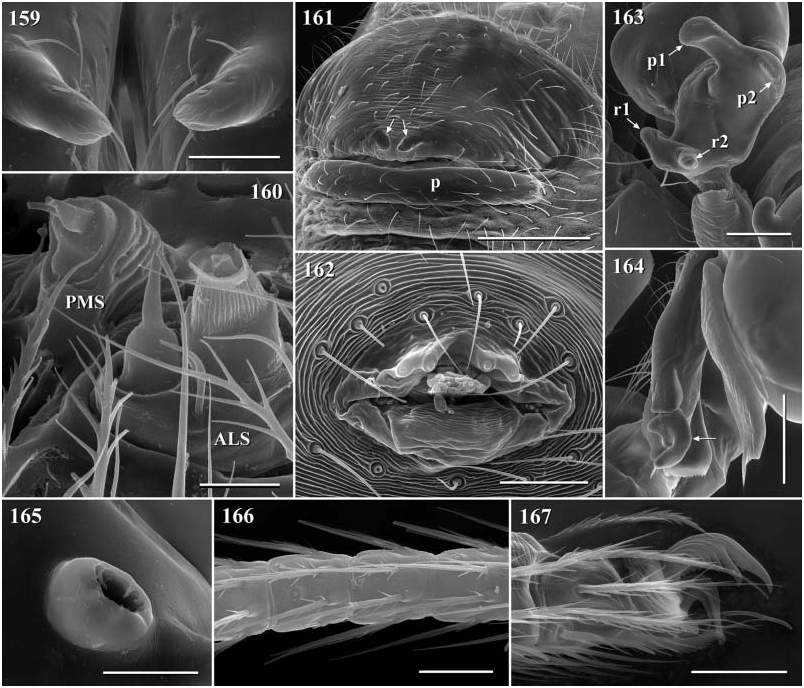
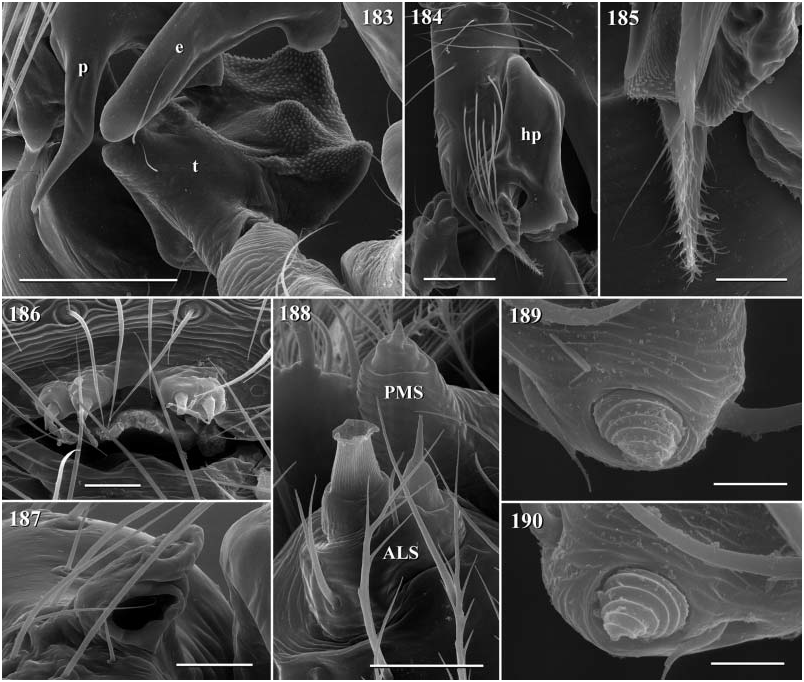
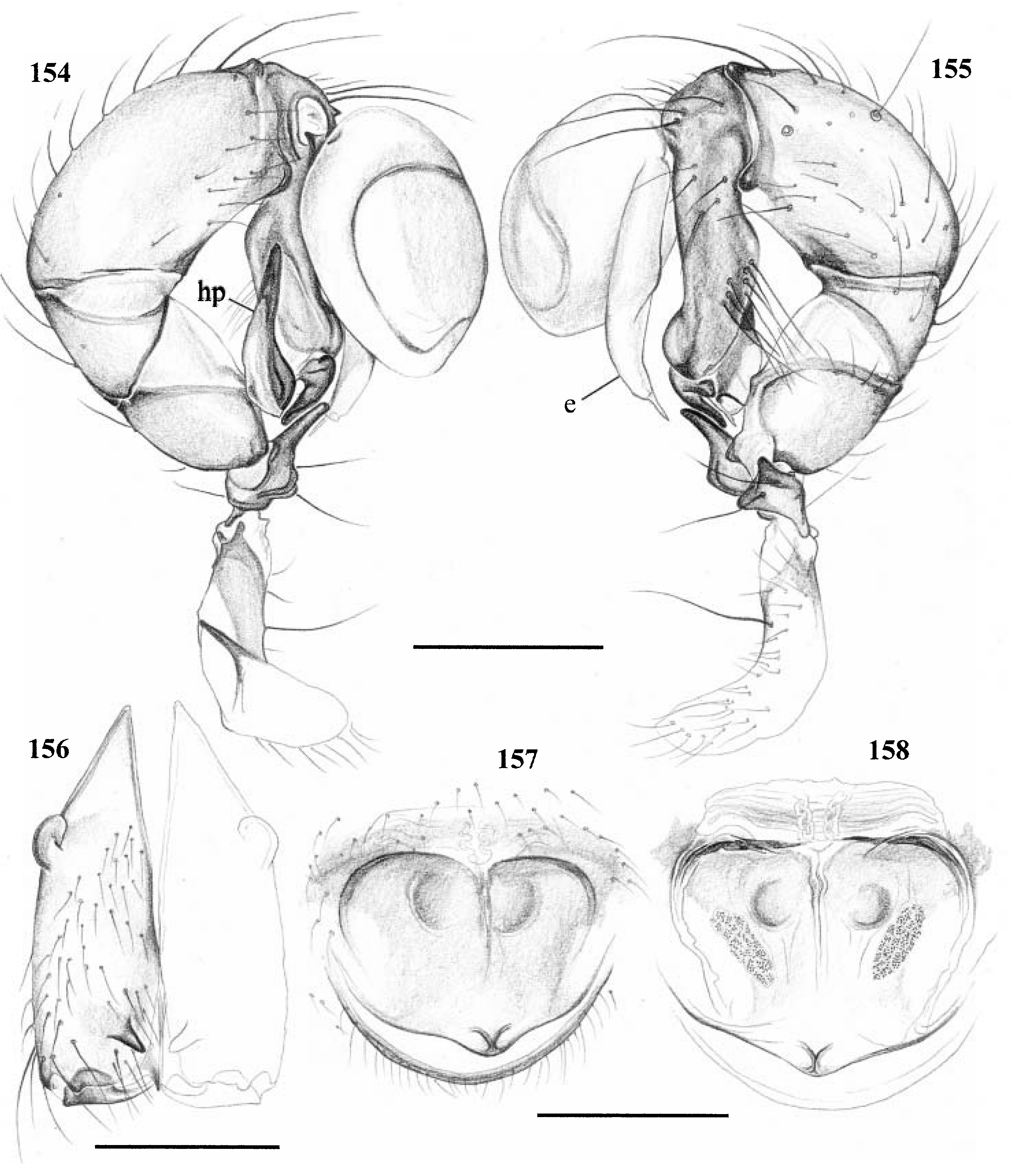
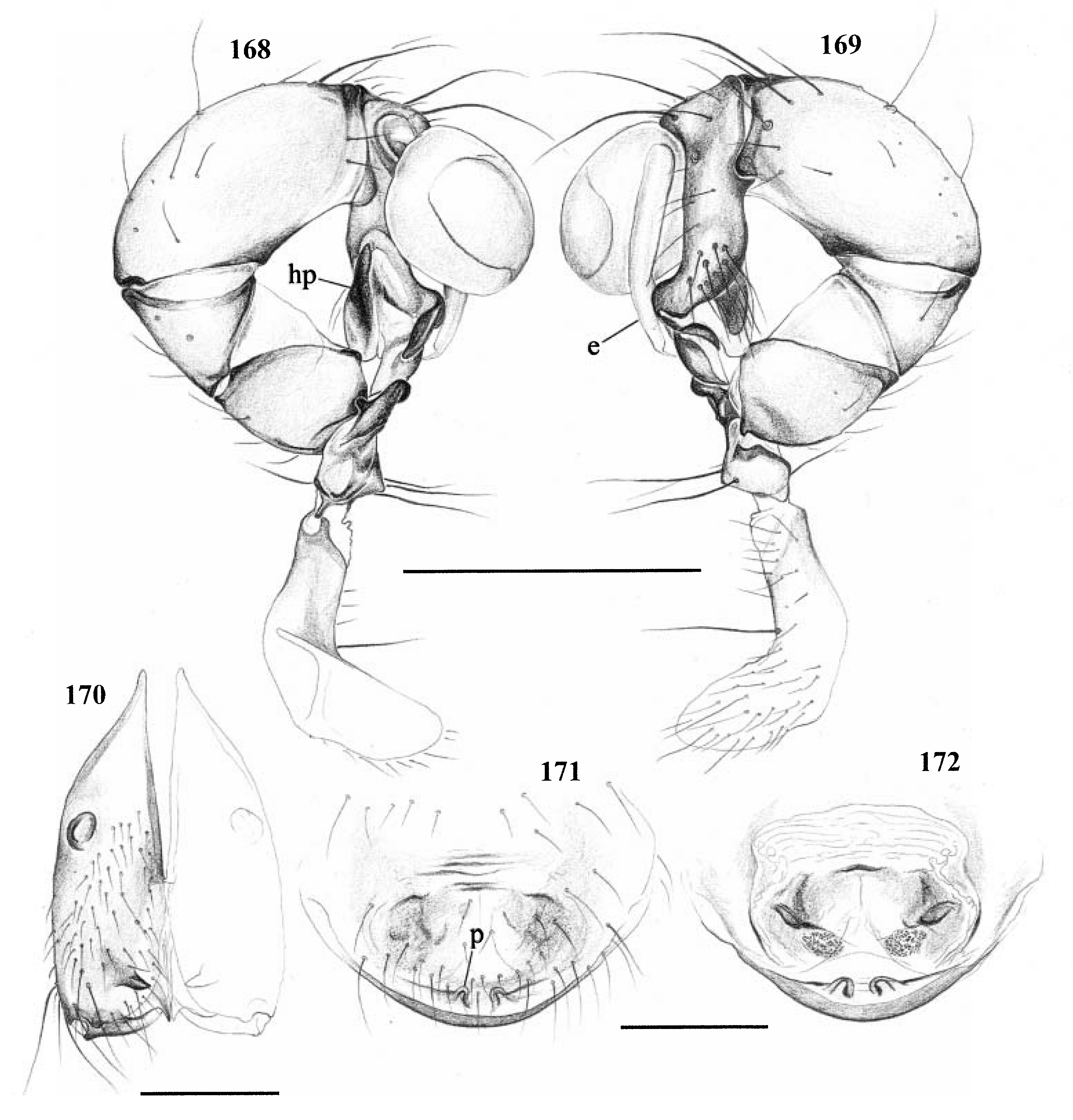
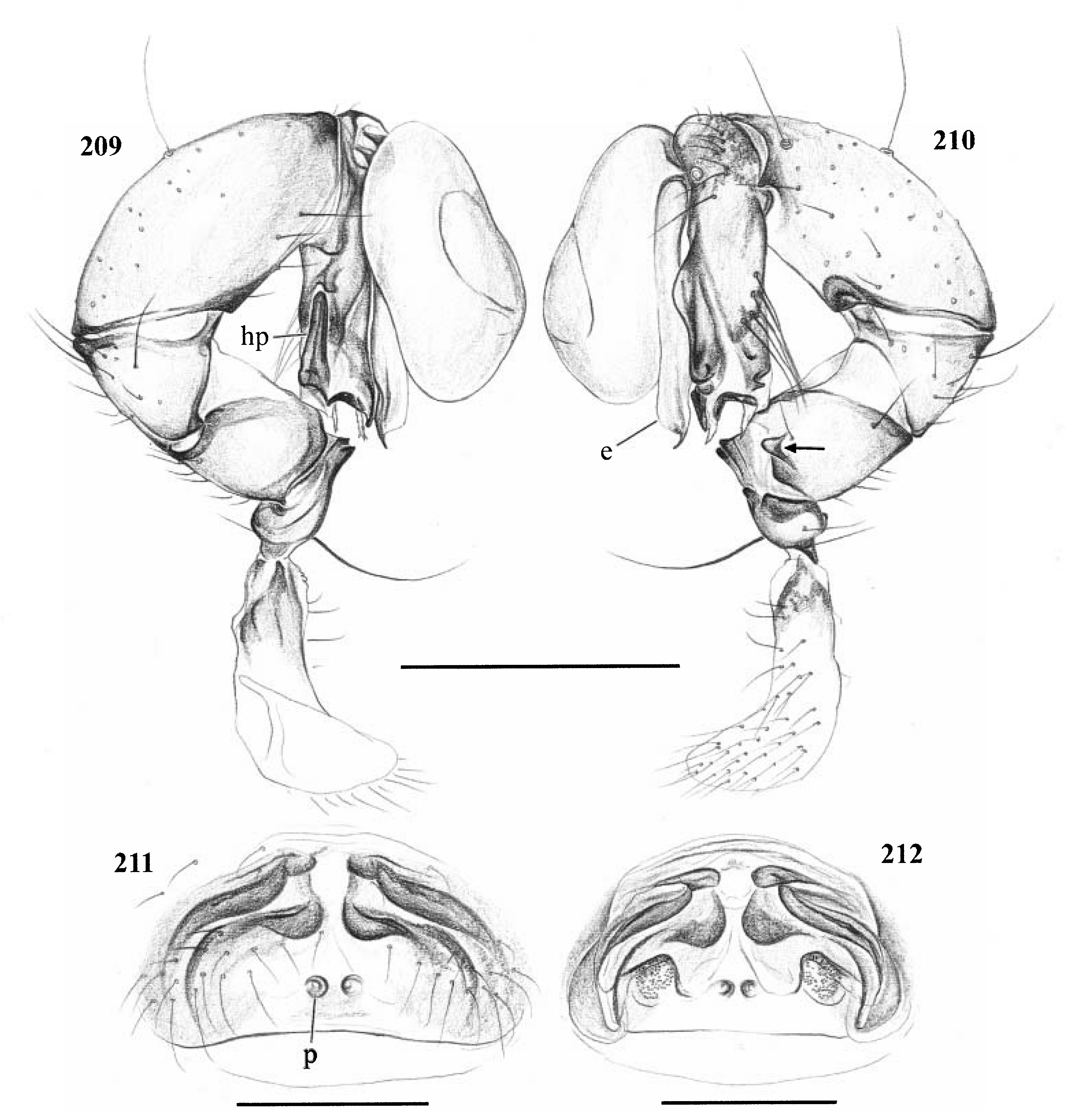
Diagnosis. Long-legged, six-eyed pholcids of medium size (total length ~ 1.5–4 mm), with oval or slightly elongate opisthosoma. Distinguished from Zatavua by lateral cheliceral apophyses not pointing backwards ( Figs 136 View Figures 131–138 , 179 View Figures 177–182 , 193 View Figures 191–197 and 201 View Figures 198–203 ), absence of notch on cymbium, wide distance between eye triads (PME- PME> 230% of PME diameter, vs. <180% in Zatavua ) and by reduction of spigots on ALS to basic set of two ( Figs 148, 149 View Figures 147–153 , 160 View Figures 159–167 and 188 View Figures 183–190 ). Distinguished from other related genera by shift of tibia-tarsus joints, resulting in dorsal position of prolateral joint and ventral position of retrolateral joint ( Figs 135 View Figures 131–138 , 155 View Figures 154–158 , 169 View Figures 168–172 , 178 View Figures 177–182 , 192 View Figures 191–197 , 199 View Figures 198–203 and 205 View Figures 204–208 ).
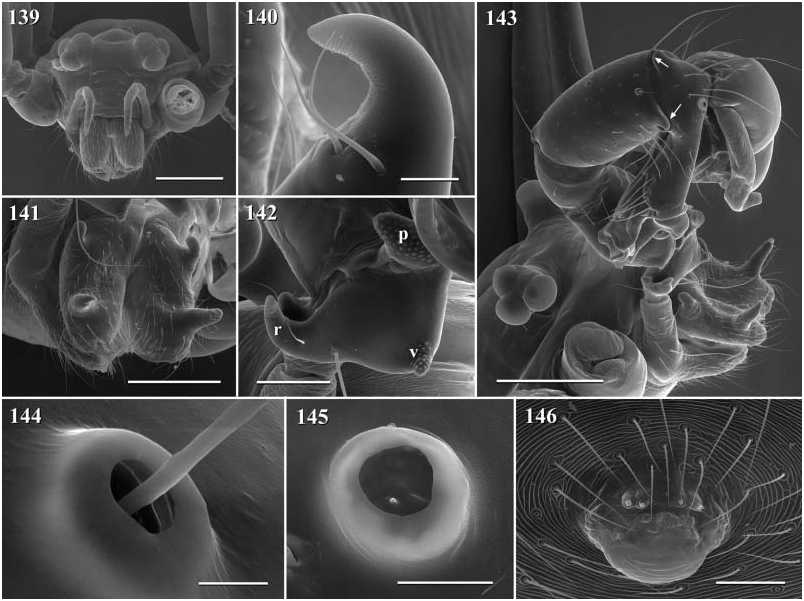
Description. Total length in males about 1.5–3.5 mm. Carapace oval or roundish, thoracic groove present but often very shallow and restricted to frontal half, often with dark pattern that may be distinctive for species. Six eyes in two triads, on moderately elevated ocular area. Distance PME-ALE small (~30–50% of PME diameter), distance PME-PME large (~230– 500% of PME diameter); no trace of AME. Clypeus never modified. Male chelicerae with pair of simple lateral apophyses and frontal apophyses in variable position and shape; the latter in some species with modified hairs imbedded in tips ( P. nampoinai , manantenina ; Figs 189, 190 View Figures 183–190 and 195 View Figures 191–197 ). Male palps small to very large in relation to overall size (compare Figs 109 and 113 View Figures 102–121 ); coxa without apophysis, in some species with ventral hump; trochanter complex, with several sclerotized apophyses; femur very short, in some species with dorso-distal apophysis (arrows in Figs 178 View Figures 177–182 , 192 View Figures 191–197 and 199 View Figures 198–203 ); patella triangular in lateral view; tibia cylindrical, not highly expanded, with two trichobothria; tibia-cymbium joints distinctively shifted, resulting in dorsal position of prolateral joint and ventral position of retrolateral joint ( Figs 135 View Figures 131–138 , 155 View Figures 154–158 , 169 View Figures 168–172 , 178 View Figures 177–182 , 192 View Figures 191–197 , 199 View Figures 198–203 and 205 View Figures 204–208 ); procursus usually complex, with cup-shaped tarsal organ (e.g. Figs 145 View Figures 139–146 , 150 View Figures 147–153 and 165 View Figures 159–167 ), in proximal position (e.g. Figs 135 View Figures 131–138 and 174 View Figures 173–176 ), with hinged process prolatero-ventrally (‘hp’ in Figs 154 View Figures 154–158 , 168 View Figures 168–172 and 209 View Figures 209–212 ); bulb consisting of proximal globular part and membranous embolus that is sometimes provided with distal transparent spine. Legs usually long (leg 1 about 10–13 ¥ body length), medium-thin (tibia 1 L/d ~50–90), leg 1 always longest, legs 2 and 4 about same length, leg 3 shortest. Legs sometimes with macrosetae ventrally on femora (only femora 1, rarely also 2), with few vertical hairs, without curved hairs; retrolateral trichobothrium of tibia 1 at 7–12%. Prolateral trichobothrium missing on tibiae 1, present in all others. Tarsus 1 with over 20 pseudosegments, but only ~10–20 distal pseudosegments easily visible in dissecting microscope. Opisthosoma either oval or slightly elongate, rarely with slight posterior elongation over spinnerets ( Figs 105 and 107 View Figures 102–121 ). Male gonopore with four epiandrous spigots in all species examined ( P. betsileo , coddingtoni , manantenina ; Figs 146 View Figures 139–146 , 162 View Figures 159–167 and 186 View Figures 183–190 ); ALS with only basic set of two spigots (examined: P. betsileo , coddingtoni , ralamboi , manantenina , nampoinai , scharffi , rothorum , marojejy , rabeariveloi ; Figs 148, 149 View Figures 147–153 , 160 View Figures 159–167 and 188 View Figures 183–190 ); other spinnerets typical for family ( Fig. 147 View Figures 147–153 ; cf. Huber, 2000).
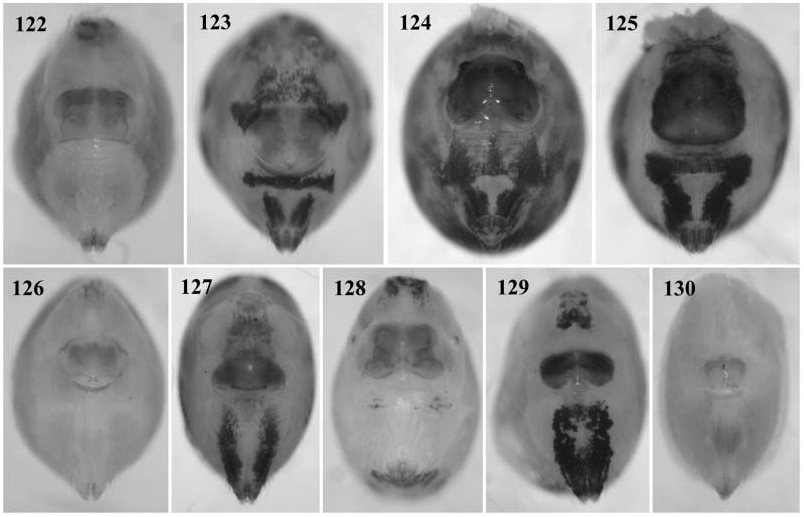
Sexual dimorphism slight. Colour pattern often more variable in females than in males. Female legs never with spines. Epigynum shape very variable ( Figs 122–130 View Figures 122–130 ), often with pair of pockets.
Monophyly. The synapomorphy supporting this genus is the shift of the tibia-cymbium joints (see Description above).
Generic relationships. As discussed above, the sister group of Paramicromerys is not resolved in any of the 12 most parsimonious cladograms obtained. Instead, Paramicromerys is part of a polytomy that always includes East African ‘ Spermophora ’, along with either Spermophorides or Metagonia and Micromerys (Appendix 5). All these genera share a hinged process on the procursus that is here interpreted as homologous, and the reduction of the spigots on the ALS to the basic set of two.
Specific relationships. A close relationship between P. nampoinai and manantenina is supported by several synapomorphies: modified hairs on the tips of the cheliceral apophyses, short side branch on embolus, sclerotized cone-shaped structure proximally on procursus ( Figs 178 View Figures 177–182 and 192 View Figures 191–197 ) that carries the tarsal organ ( Fig. 187 View Figures 183–190 ). Both species share with P. scharffi a proximo-dorsal apophysis on the male palpal femur (arrows in Figs 178 View Figures 177–182 , 192 View Figures 191–197 and 199 View Figures 198–203 ). A second speciesgroup is characterized by a transparent spine distally on the embolus [ P. coddingtoni and the putatively close relatives combesi (Millot) , ralamboi , quinteri , mahira ], but this might be a plesiomorphy. Both species groups share the spines ventrally on the frontal male femora [unknown in P. quinteri , mahira , and combesi (Millot) ]. The type species P. madagascariensis (Simon) and P. betsileo have almost identical palps (compare Figs 131 and 135 View Figures 131–138 ) and are therefore considered closely related. The other species included ( P. rothorum , marojejy , rabeariveloi ) all share the shift of the tibia-cymbium joints, but their exact position is unclear. Finally, P. megaceros (Millot) is tentatively assigned to Paramicromerys because it shares the wide distance between the eye triads and the simple lateral cheliceral apophyses, but it may be misplaced.
Natural history. No species has ever been studied in any detail, and little can be inferred from notes on the collection labels. In contrast to Zatavua , only one known species is cavernicole [the tentatively assigned P. megaceros (Millot) ]. Paramicromerys betsileo has been collected abundantly on low foliage and saplings, by fogging of dead leaves on fallen trees, on roadside vegetation, and by beating low vegetation (often together with ‘Spermophora’ ranomafana n. sp.).
Distribution. Known from Madagascar only. Most records are from just three localities (Map 2), but the high diversity at each of these localities suggests that the genus has a much wider distribution. No species has been collected at two localities more than a few kilometres apart. This is especially striking in the north, where two locations less than 200 km apart (Montagne d’Ambre and Marojejy Reserve ) yielded four and five species, respectively (one species from Marojejy is not described herein), with no overlap.
Composition. The genus includes a total of 14 described species. Of these, two were described by Millot (1946), and are not treated below ( P. combesi , megaceros ). The remaining 12 species [11 new species and P. madagascariensis (Simon) ], are described
Map 2. Known distribution of Paramicromerys and of Malagasy ‘ Spermophora ’ species.
below. The collections studied contain three additional new species that are not described due to their poor state of preservation. Considering the facts that (1) most or all species seem to have small distributional ranges (see above) and that (2) all areas that were subjected to intense collecting yielded between four and five species, it seems reasonable to expect several dozen additional species not yet discovered.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Paramicromerys
| Huber, Bernhard A. 2003 |
Paramicromerys
| Millot J 1946: 145 |