
Ascalabos voithii Graf
|
publication ID |
https://doi.org/10.5194/fr-19-31-2016 |
|
DOI |
https://doi.org/10.5281/zenodo.11045888 |
|
persistent identifier |
https://treatment.plazi.org/id/03F68791-BA33-2F11-FFFF-FD43FF55D291 |
|
treatment provided by |
Felipe |
|
scientific name |
Ascalabos voithii Graf |
| status |
|
Ascalabos voithii Graf zu Münster, 1839b
Figures 14–19 View Figure 14 View Figure 15 View Figure 16 View Figure 17 View Figure 18 View Figure 19
1839b Ascalabos Voithii – Graf zu Münster, p. 112, pl. XII, fig. 5.
1843 Leptolepis Voithii – Agassiz, p. 131, 165, 295, pl. LXI, figs. 2–3.
1843 Leptolepis polyspondylus – Agassiz, p. 133, pl. LXI, figs. 7–8.
1843 Leptolepis paucispondylous (nomen nudum) – Agassiz, p. 134.
1848 Leptolepis Voithii – Giebel, p. 143.
1848 Tharsis radiatus – Giebel, p. 146.
1863 Leptolepis Voithii, Wagner , p. 131, 134.
1895 Leptolepis voithii – Woodward, p. 512.
1974 Ascalabos voithi – Nybelin, 173–180, text-figs. 35, 36, pl. XXIX, figs. 1–5, pl. XXX.
1975a Leptolepis ( Ascalabos) voithi – Taverne, 233–243, figs. 1–6.
1977 Ascalabos voithii – Patterson and Rosen, 151–152, figs. 52–53.
1991 Ascalabos voithi – Arratia, 268–271, fig. 9.
1997 Ascalabos voithii – Arratia, 31–39, figs. 14–21.
Neotype: The holotype is lost. Nybelin (1974) designated as neotype a specimen identified as Eichstätt I (currently JME SOS 537 ).
Additional specimens: CM 9491. CMMH 9491. JME SOS 2362, JME SOS 2364, JME SOS 2365, JME SOS 2458, JME SOS 2483, JME SOS 2496, JME SOS 2497, JME SOS 2886 (peels of scales), and many other specimens deposited at the JME. NHM 3672, NHM 3673a, NHM 37062, and NHM 37080. SHL, collection of H. Leich (Bochum, Germany) currently deposited in Tierpark und Fossilium Bochum (Bochum). SMNH P5683. TM 6651, TM 10307, and TM 10325. Additionally, one uncatalogued specimen belonging to U. Eller (Dümpelfeld, Germany) was included because of its excellent preservation.
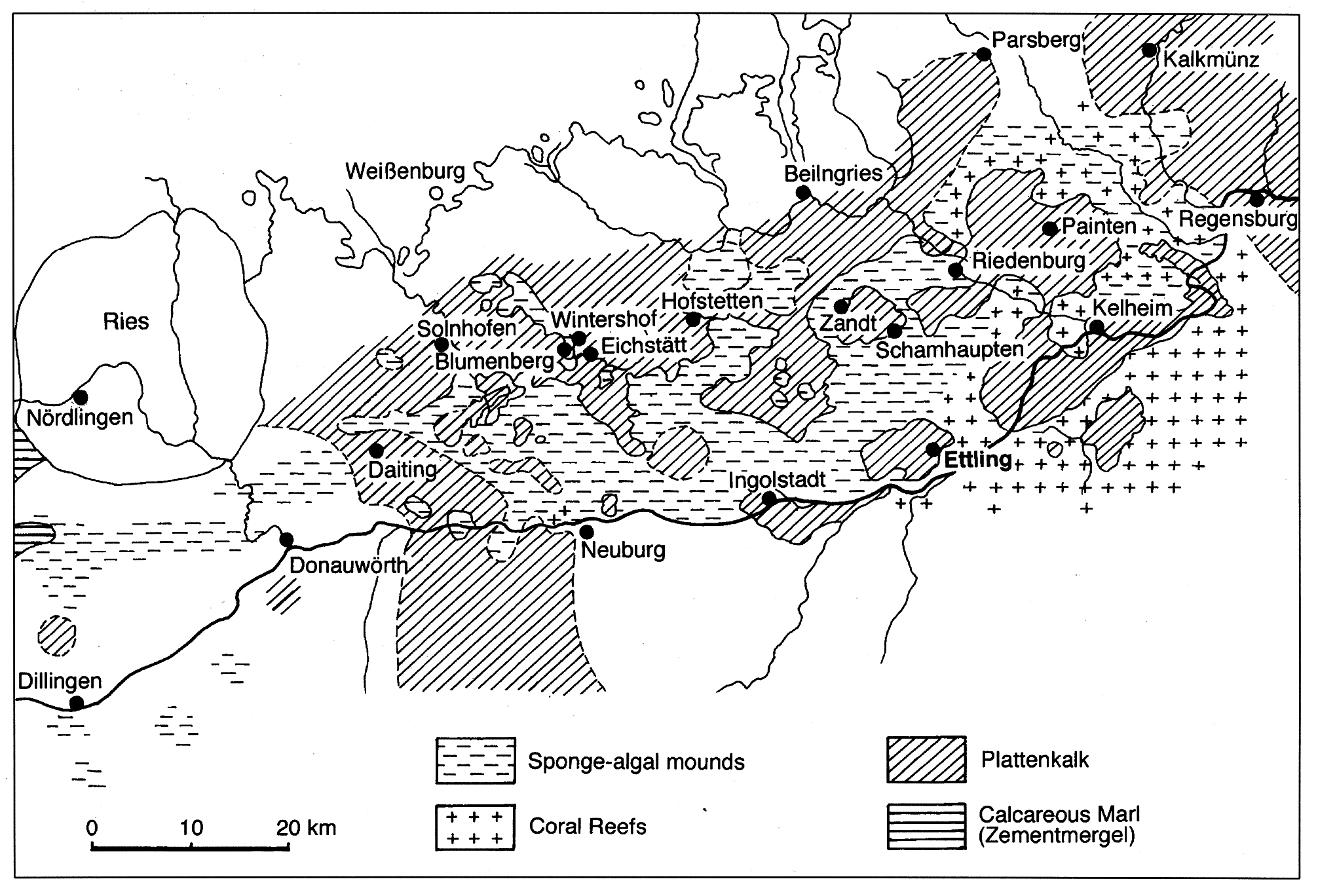
Locality and geological time: Blumenberg, Eichstätt, Hofstetten, Kelheim, and Wintershof ( Fig. 1 View Figure 1 ) in Bavaria, southern Germany. Upper Jurassic, Tithonian ( Meyer and Schmidt-Kaler, 1989, 1990; Schweigert, 2015). Ascalabos voithii has also been cited from Cerin ( France; de Saint-Seine, 1949; Wenz et al., 1993), which is interpreted as Kimmeridgian in age. This material is currently under revision.
Description
A description of Ascalabos voithii can be found in Arratia (1997). Here, I only describe and illustrate additional information after of a re-study of the neotype and other specimens. I describe some characters that add new interpretations or a more complete understanding of certain structures. Some are compared with the structures in Ebertichthys n. gen.
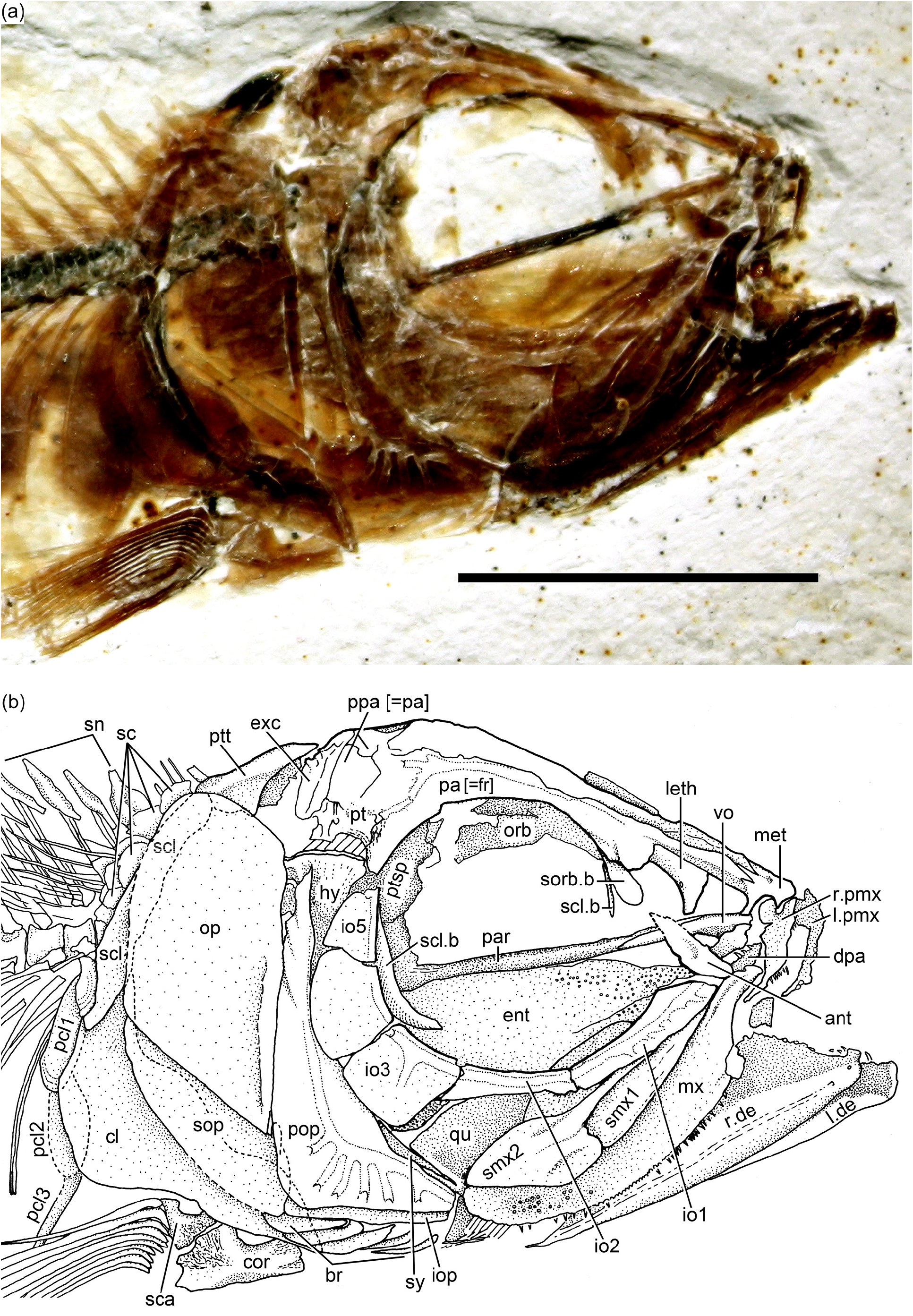
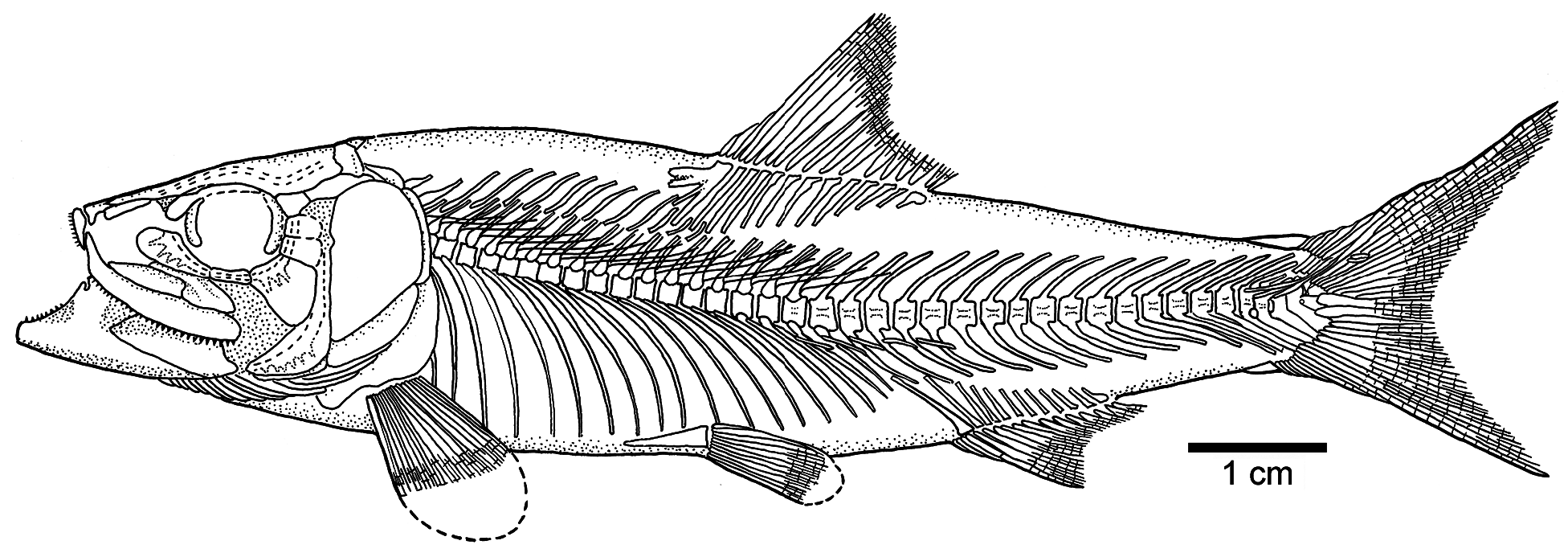
General description: Ascalabos voithii has a head proportionally large, about 30 % of standard length; however, the head in Ebertichthys is comparative larger, about 34 % of SL. The diameter of the eye is large, 22 to 25 % of the head length ( Figs. 14b View Figure 14 and 15b View Figure 15 ), whereas the eye in Ebertichthys ettlingensis is even larger, 34 to 36 % of the head length. The insertion of the dorsal fin ( Figs. 14a View Figure 14 , 15a View Figure 15 , and 16 View Figure 16 ) is above the level of the insertion of the pelvic fins, about 52 to 58 % of SL. In contrast, the insertion of the dorsal fin is posterior to that of the pelvic fins in Ebertichthys ettlingensis .
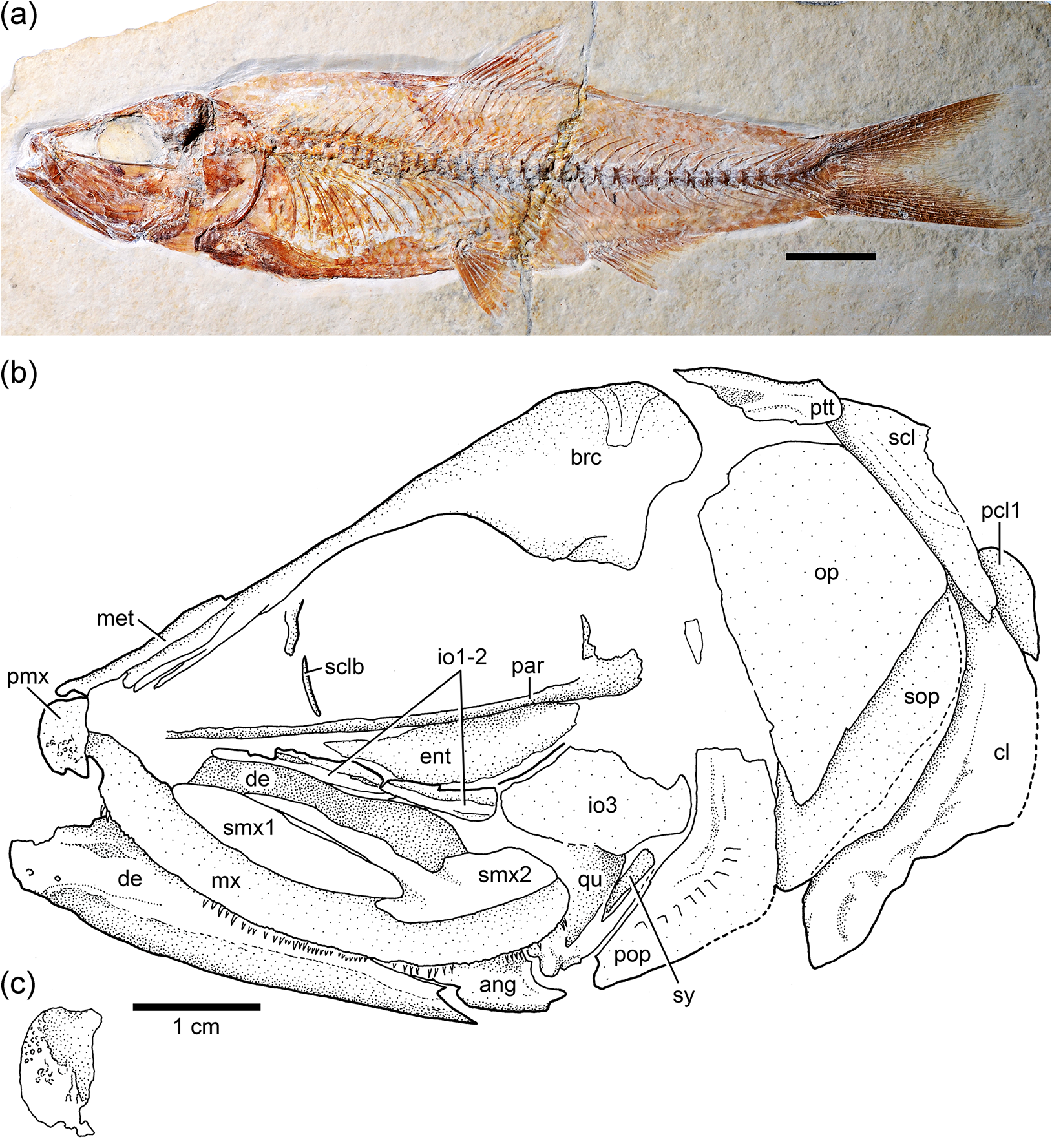
Cranial bones: The cranial bones are unornamented with the exception of the premaxilla, which bears some small, tubercle-like ornaments ( Fig. 14c View Figure 14 ) covered with a thin layer of ganoine in the neotype. The braincase of the neotype is poorly preserved and does not add new information.
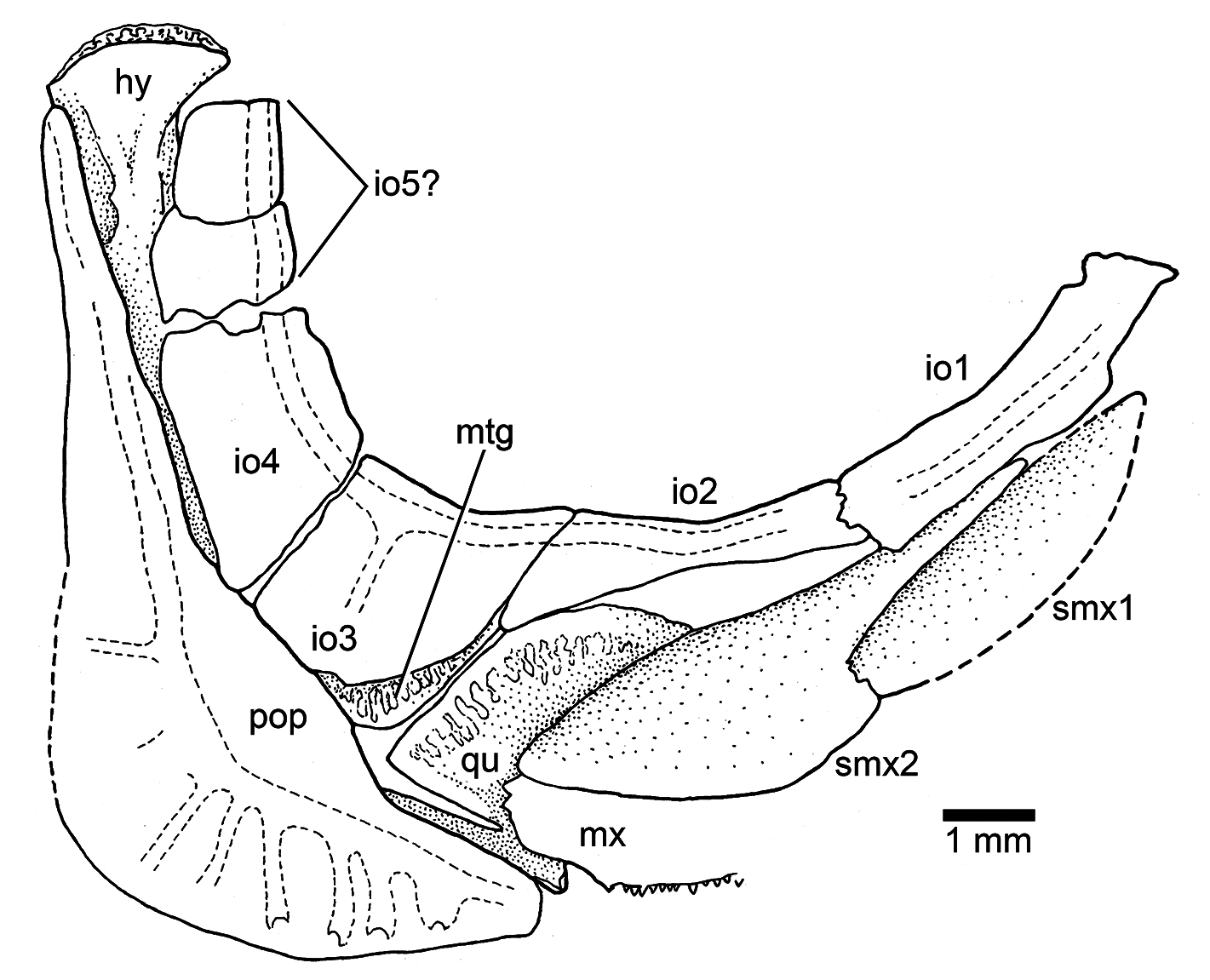
Circumorbital series: The supraorbital is incompletely preserved in most specimens or not preserved at all. It has an expanded anteroventral region, similar to that found in Ebertichthys ettlingensis . Infraorbital 1 is an elongate bone that joins with an elongate infraorbital 2 that is not as long as that of Ebertichthys (compare Figs. 3b View Figure 3 , 4a View Figure 4 , and 5 View Figure 5 with Fig. 14b View Figure 14 ). Infraorbital 3 is a small bone at the posteroventral corner of the orbit ( Fig. 14b View Figure 14 ). Because of its reduced size, a significant part of the quadrate is exposed. Although dorsoposterior infraorbitals 4 and 5 as the dorsal part of the preopercle are not preserved in the neotype, space left indicates that the region between the inner margin of infraorbitals 3–5 and the anterior margin of the preopercule was narrow.
Upper jaw: It contains the premaxilla, maxilla, and two supramaxillae ( Fig. 14b View Figure 14 ), which are all preserved in the neotype. The re-study of the neotype confirms that the premaxilla has a short and small ascendent process as in the premaxilla in Ebertichthys n. gen. However, the articulatory anterior process of the maxilla is not as long as that in Ebertichthys , so that the posterior blade of the premaxilla is shorter. The maxilla is long and partially covers the anterior part of quadrate. The two supramaxillae of the neotype are very similar to that of Ebertichthys ettlingensis (compare Figs. 3b View Figure 3 and 12b View Figure 12 ). Supramaxilla 2 is broadly expanded posteriorly, forming the main body of the bone. The anterodorsal process is very long and narrow. Supramaxilla 1 is elongate and ovoid-shaped. Both supramaxillae cover the dorsal margin of the maxillary blade.
Lower jaw: The mandible ( Fig. 14b View Figure 14 ) is strongly ossified, and although it is largely hidden by the maxilla and supramaxillae, the large and high coronoid process is visible in the neotype. The articulation of the mandible with the quadrate is placed at the posterior margin of the orbit in the neotype (specimen with almost closed mouth), which is in contrast to an anterior position in specimens with the mouth open. A “leptolepid” notch is present in the ascending dorsal margin of the dentary in specimen JME SOS 3228 . This notch is not observed in the neotype, because the maxilla covers this region. The “leptolepid” notch is narrow and partially forms the anterior wall of the massive coronoid process.
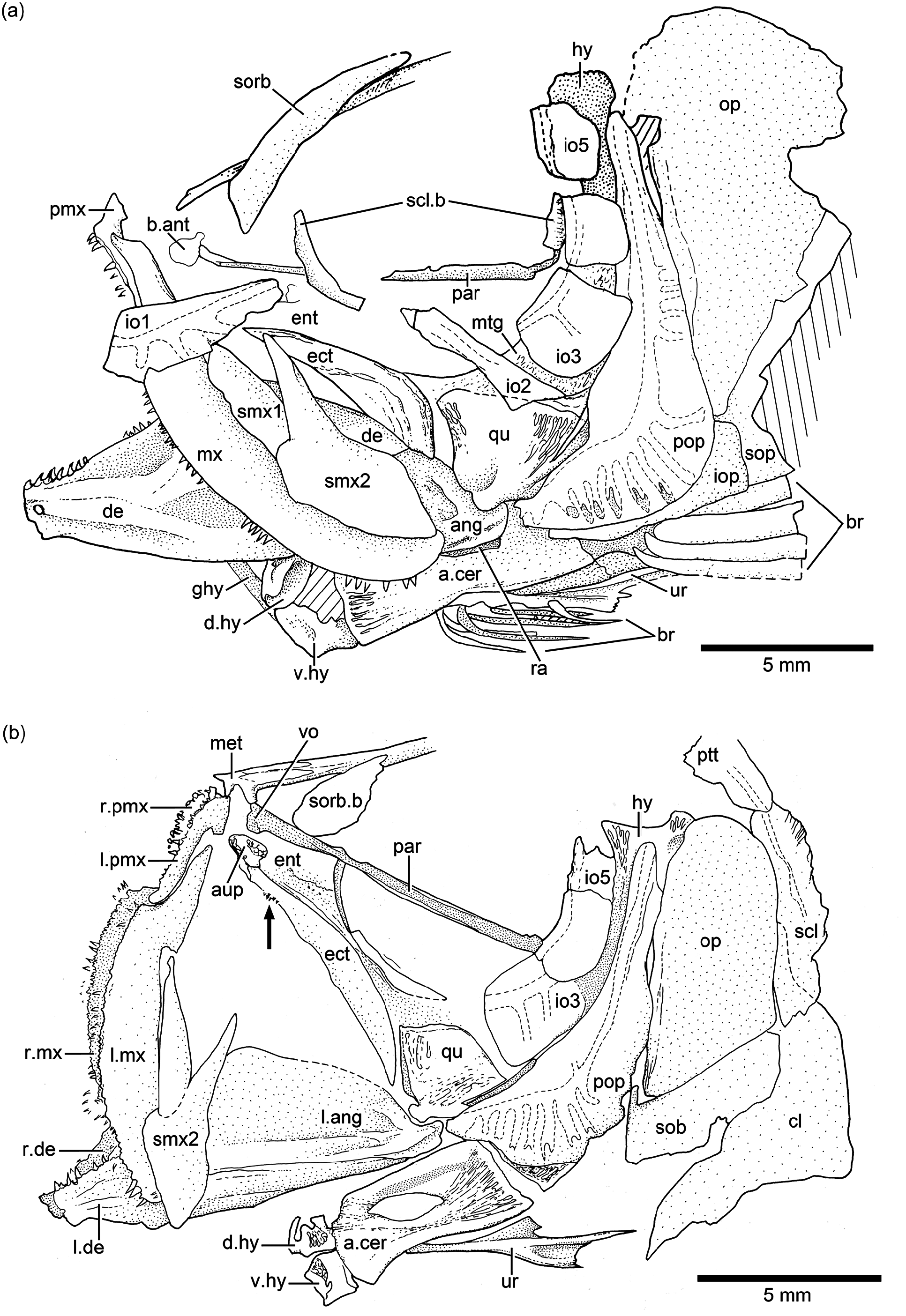
Opercular bones: The preopercle ( Figs. 14b View Figure 14 and 15b View Figure 15 ) is incompletely preserved in the neotype, and it is probably somehow displaced. The important aspect to be noted is that the bone presents a slightly rounded bony flange at its anterior margin, similar to that in Ebertichthys n. gen. (compare Figs. 3b View Figure 3 and 4a View Figure 4 with Fig. 14b View Figure 14 ). Since this feature has been observed only in a few specimens due to poor preservation, I have not considered it as a diagnostic character of the family, but this feature should be re-evaluated when more specimens of Ascalabos become available. The characteristic sensory tubules of the preopercular sensory canal are nicely preserved in the neotype ( Fig. 14b View Figure 14 ), as well as in an additional specimen illustrated in Fig. 15b View Figure 15 .
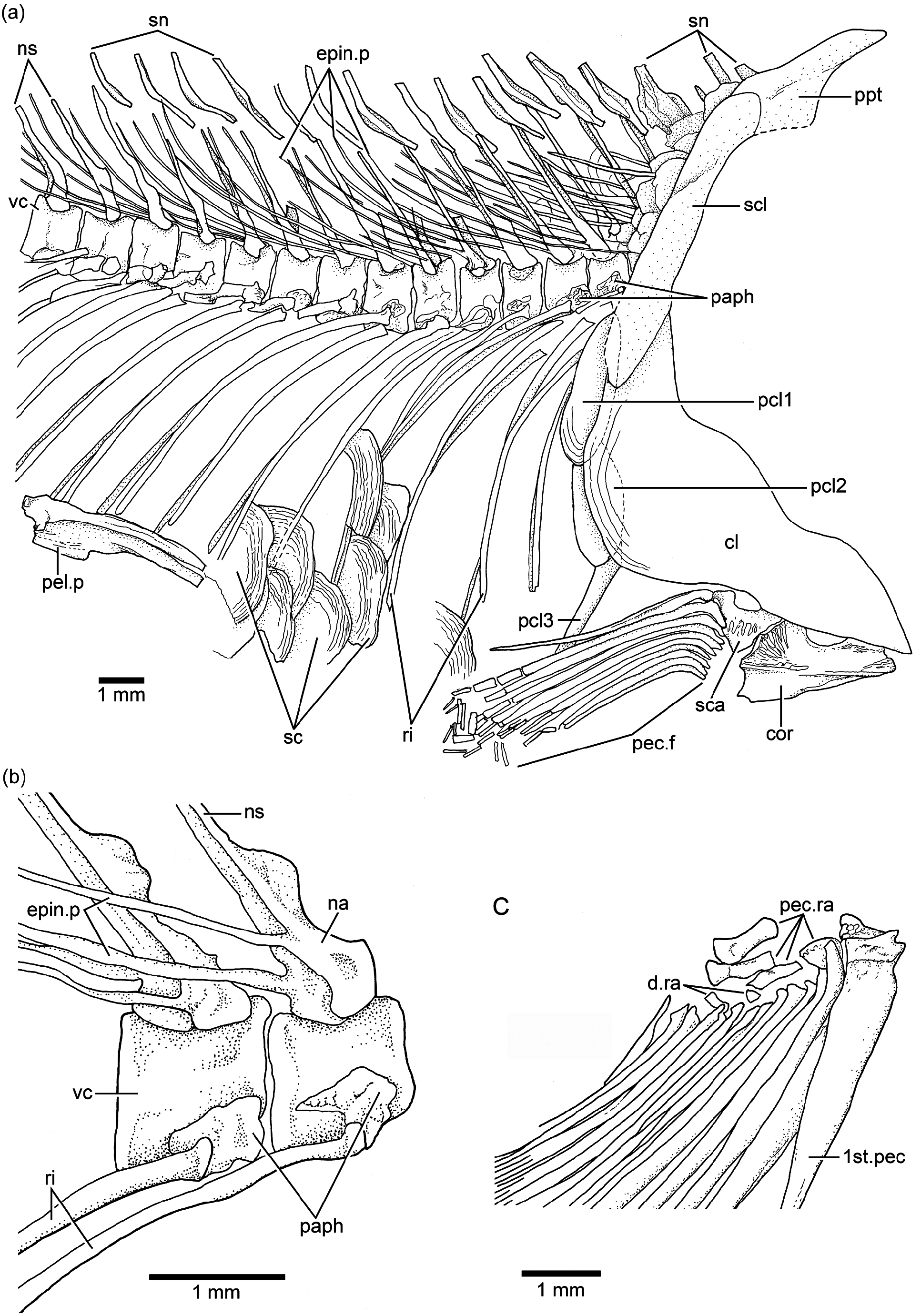
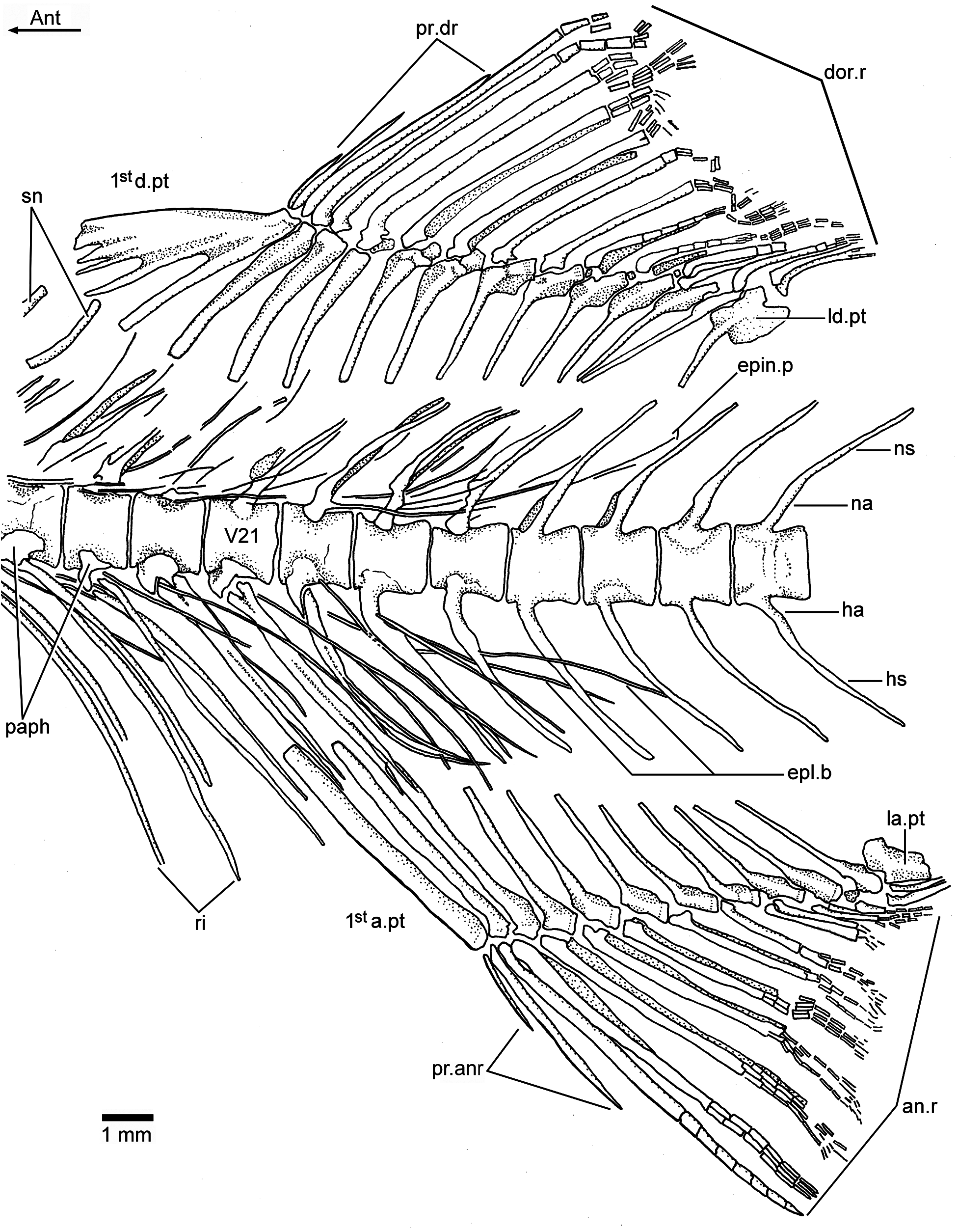
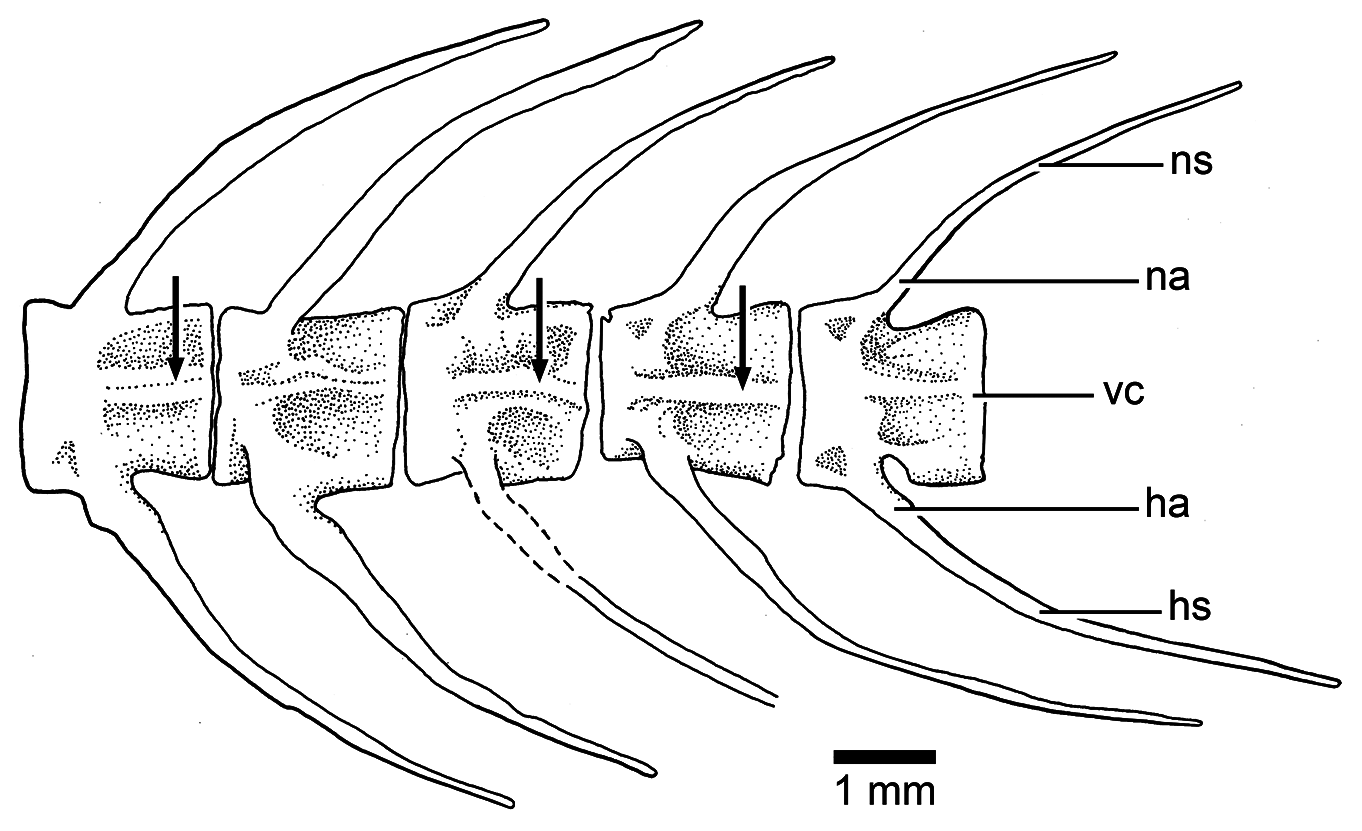
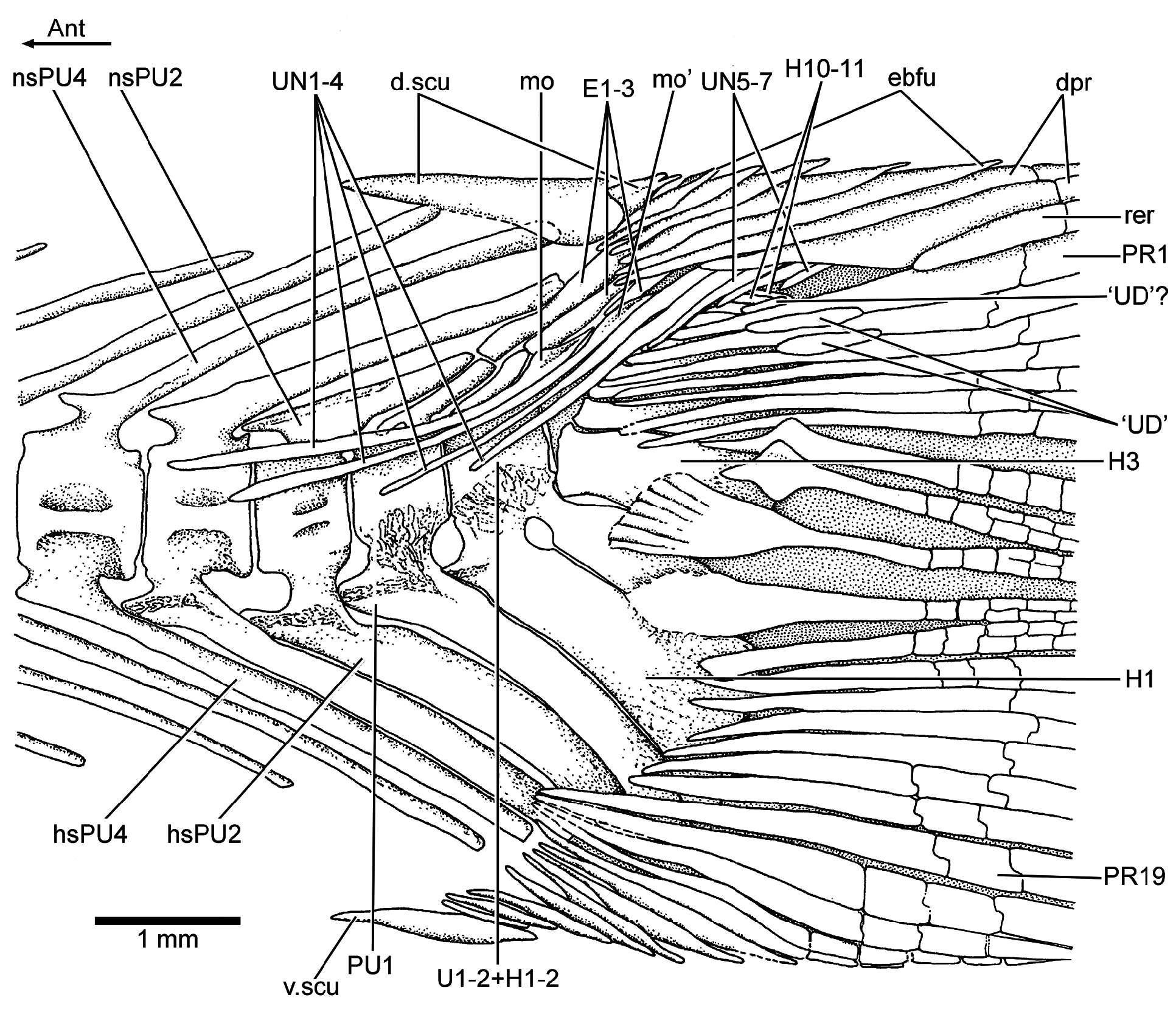
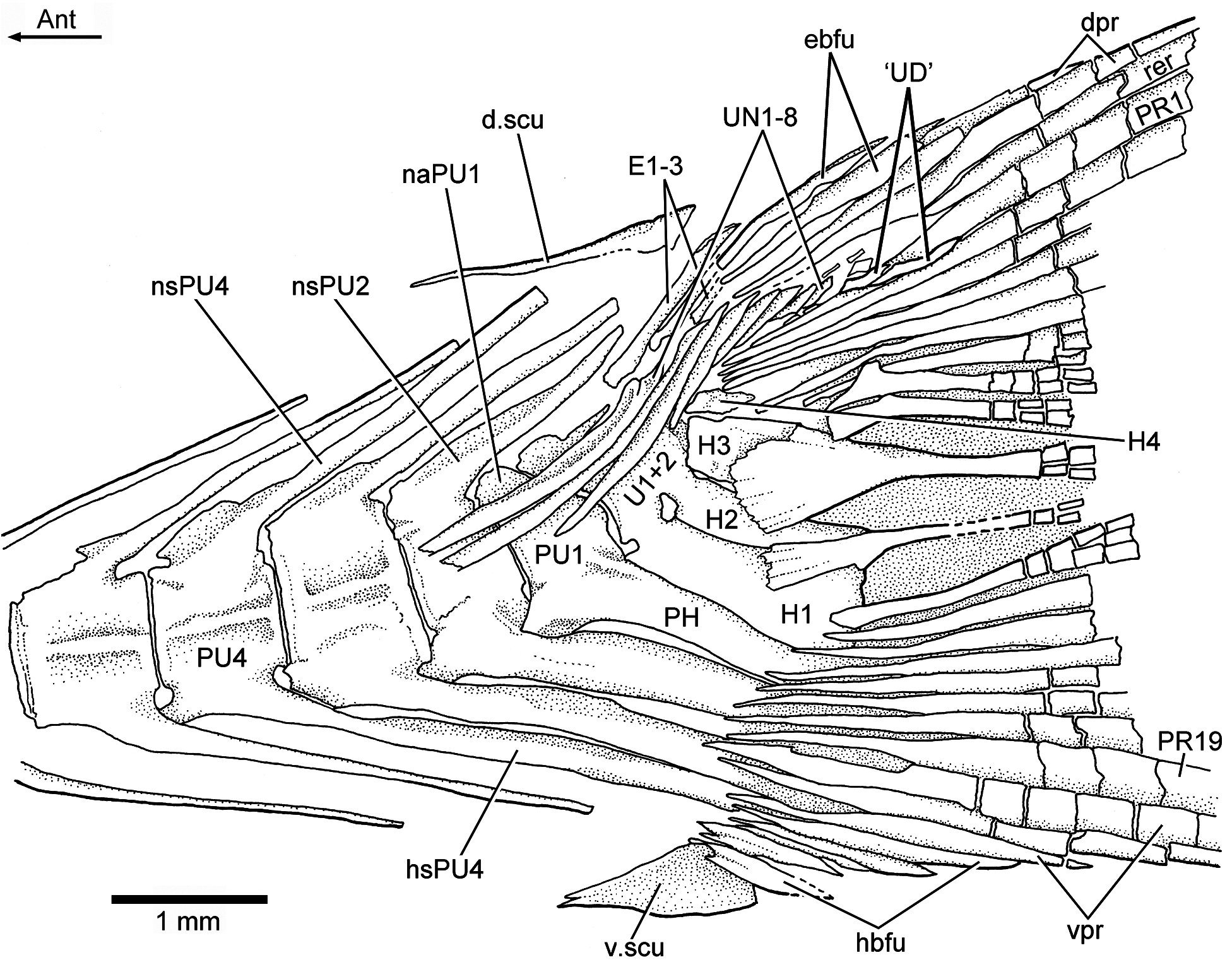
Vertebral column: A long and prominent lateral crest ( Fig. 17 View Figure 17 ) characterizes the centra of the caudal region of the vertebral column in large specimens. The prominent crest is still present on the lateral surface of preural centrum 2 ( Fig. 18 View Figure 18 ). This is very different to the condition in Ebertichthys with smooth centra. Ascalabos may have from 34 to 39 vertebrae (commonly 36), whereas in Ebertichthys the number is commonly 38 or 39. These represent the lowest counts among teleosts from the Solnhofen limestones (see Arratia and Schultze, 2015). There are 16 to 19 pairs of ribs ( Fig. 16 View Figure 16 ), whereas 18 or 19 pairs of ribs are present in Ebertichthys ettlingensis . A series of about 14 supraneurals extend between the occiput and below the first dorsal pterygiophores, whereas 16 supraneurals are commonly found in Ebertichthys , but they do not extend below the first dorsal pterygiophores.
The early formation of the centra of Ebertichthys ettlingensis ( Fig. 9 View Figure 9 ) is similar to that of Ascalabos voithii , so that the description above stands for both species. My observations (here and in Arratia, 1991, p. 257) and interpretations differ from those of Schaeffer and Patterson (1984), who interpreted the centra of young Tharsis , Ascalabos , and lycopterid osteoglossomorphs as diplospondylous, each formed by a precentrum and a postcentrum perichondrally ossified, and that the chordacentrum is then reabsorbed or suppressed in the middle of the centrum. These interpretations are not supported by my observations in young Tharsis , Ascalabos , and Ebertichthys n. gen.
Pelvic plate or basipterygium: The basipterygium of Ascalabos is characterized by its slightly triangular shape and its posterior expansion that has a short and sharp medial posterior process ( Arratia, 1997, Fig. 19c View Figure 19 ). In contrast, the basipterygium of Ebertichthys ettlingensis lacks the posterior expansion, and a medial process has not been observed.
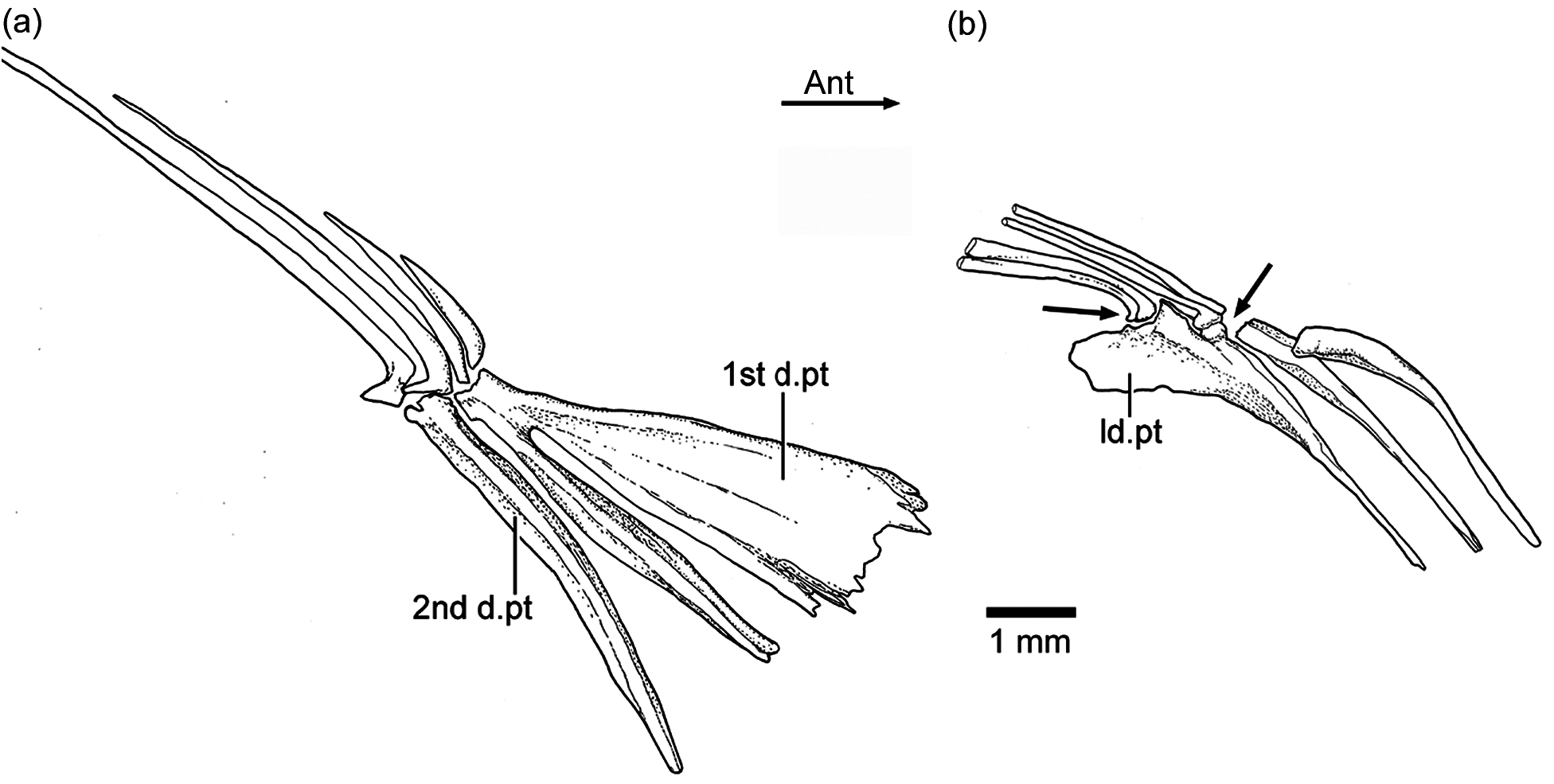
Dorsal and anal fins and their supports: The first dorsal pterygiophore is characteristically expanded and with a few anteroventral processes in Ebertichthys ettlingensis ( Figs. 7 View Figure 7 and 11a View Figure 11 ), and a similar pterygiophore ( Fig. 16 View Figure 16 ) is present in the neotype of Ascalabos . A first pterygiophore bearing only two processes (JME SOS 2886) and another partially broken one ( HL 309) were illustrated for Ascalabos by Arratia (1997, Fig. 19a–b View Figure 19 ). Currently, I interpret these bones as partially incomplete. According to the evidence gathered here, the characteristic shape of the first dorsal pterygiophore is now proposed as a diagnostic feature of the new family Ascalaboidae .
The first anal pterygiophore is long, broadly expanded and almost rectangular in outline in Ebertichthys . In contrast, the pterygiophore is thinner and more rod-like in Ascalabos as well as in other Jurassic teleosts (see Arratia, 1981, 1997, 2000).
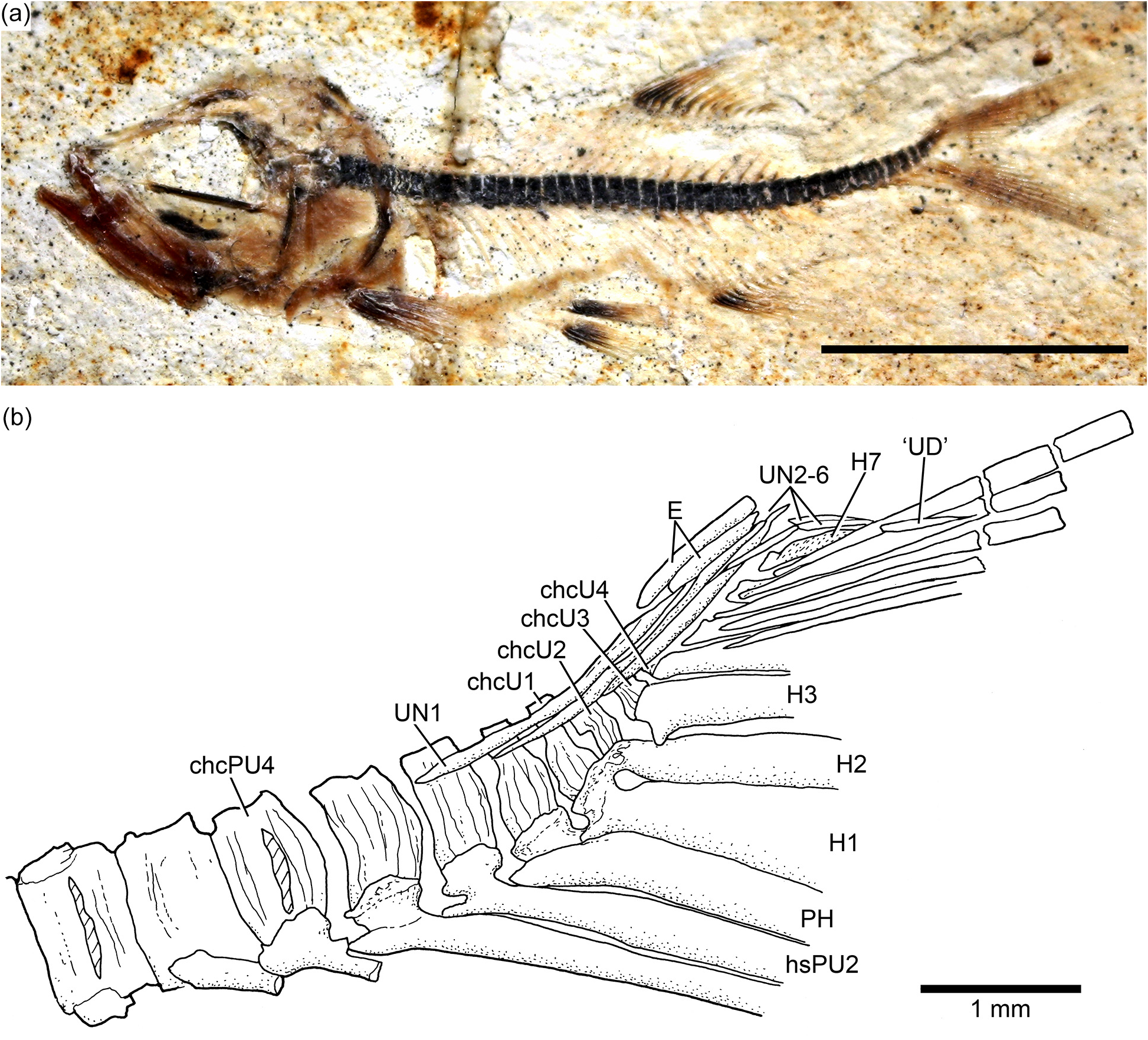
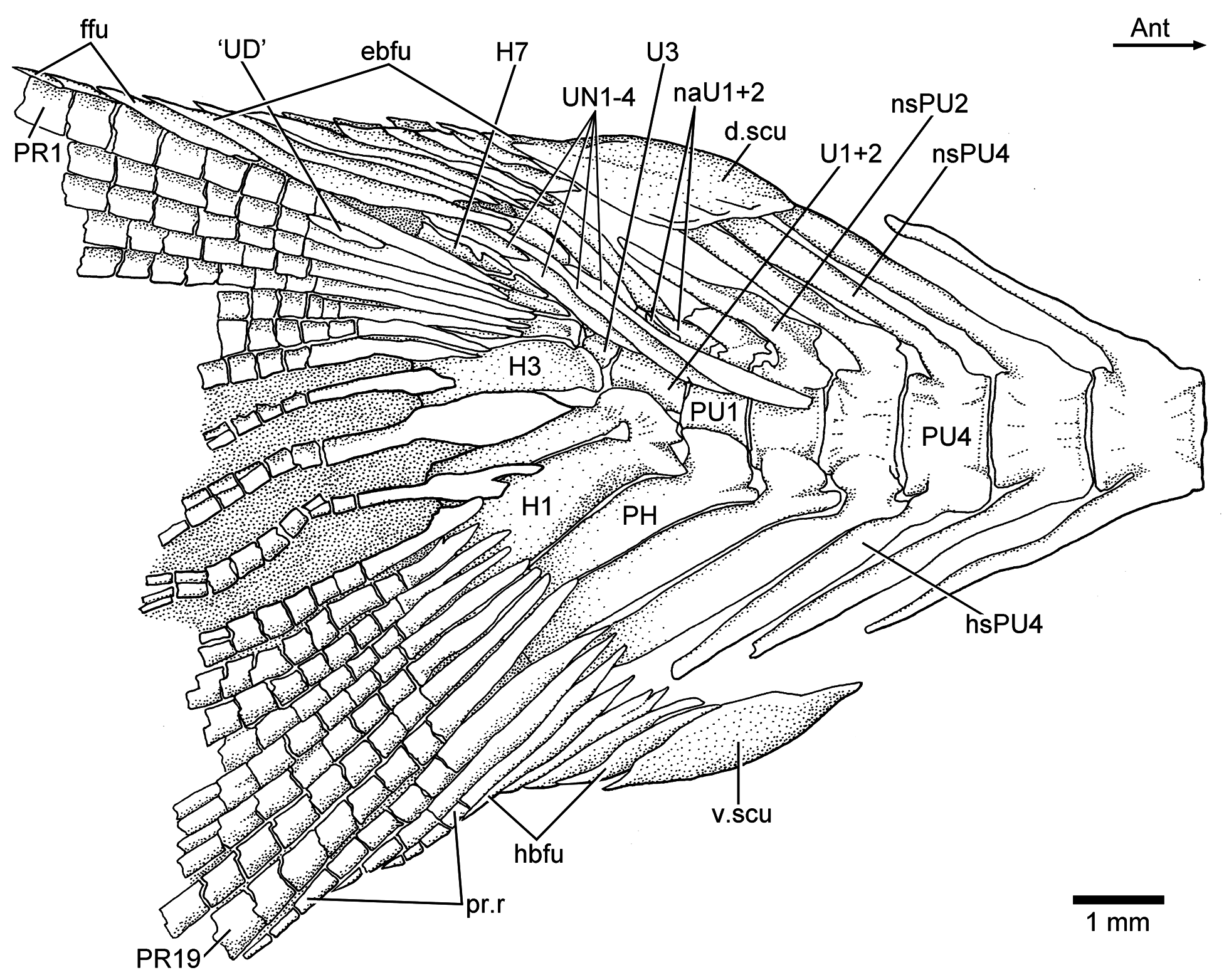
Caudal skeleton and fin: The caudal skeleton of the neotype confirms previous descriptions by Arratia (1991, 1997), but adds some new information about variability. While in some specimens the first two uroneurals extend anteriorly to the lateral surface of preural centrum 3 ( Fig. 18 View Figure 18 ), they reach preural centrum 2 in the neotype ( Fig. 19 View Figure 19 ). The neotype presents eight uroneurals, whereas in other specimens only seven have been observed, but this could be a result of poor preservation. The fourth uroneural is broken in the neotype, so that it is unclear whether it reached ural centrum 1 + 2 or not, as it does in other specimens. Uroneurals 5 to 8 reduce in size caudally. The neural spine of preural centrum 2 is slightly shorter than that of preural centrum 3, which is broken distally. The neural arch of preural centrum 1 is incompletely preserved.
There is an incomplete series of epaxial basal fulcra, two segmented procurrent rays, one epaxial rudimentary ray, 19 principal caudal rays, three hypaxial procurrent rays, and five hypaxial basal fulcra in the neotype ( Fig. 19 View Figure 19 ). Fringing fulcra are absent. In contrast, other specimens have five or six epaxial basal fulcra and seven hypaxial basal fulcra ( Fig. 18 View Figure 18 ). Dorsal and ventral scutes precede the epaxial and hypaxial basal fulcra, respectively.
Scales: The scales of Ascalabos are incompletely known, because it is difficult to observe isolated scales. The strongly imbricate scales are usually preserved in situ. They are slightly rectangular, with incomplete circuli in the anterior field and with transverse lines that do not completely cross the middle field of the scale. In contrast, Ebertichthys n. gen. has large, slightly oval scales ( Fig. 6b View Figure 6 ) with few circuli in the anterior field. No transverse lines have been observed.

| CM |
Chongqing Museum |
| SMNH |
Department of Paleozoology, Swedish Museum of Natural History |
| TM |
Teylers Museum, Paleontologische |
| HL |
Houghton Lake Wildlife Research Station |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.