
Allorapisma, Makarkin, Vladimir N. & Archibald, Bruce, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.186898 |
|
DOI |
https://doi.org/10.5281/zenodo.6226718 |
|
persistent identifier |
https://treatment.plazi.org/id/03EFA45A-FFAF-2422-FF3D-1988FE2E9D7F |
|
treatment provided by |
Plazi |
|
scientific name |
Allorapisma |
| status |
gen. nov. |
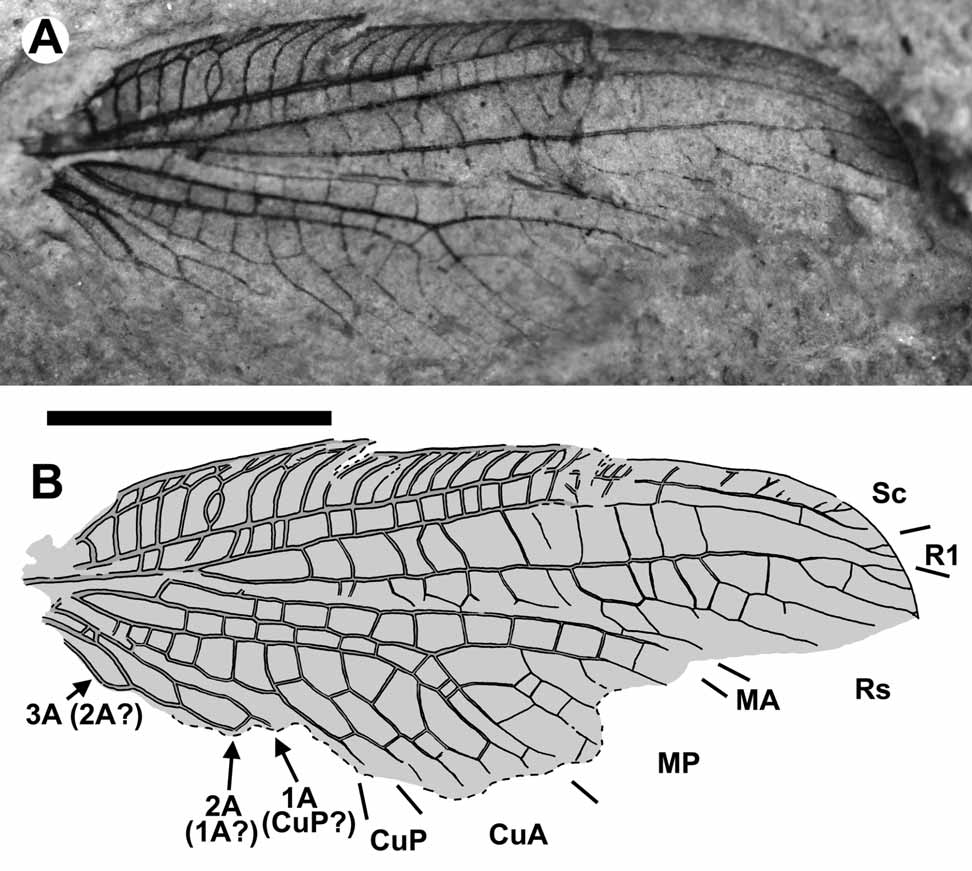
Genus Allorapisma View in CoL gen. nov.
Etymology. Allo- (from the Greek allo, other) + - rapisma (from Rapisma , a genus-group name), in reference to its venational similarity to the genus Rapisma Navás. Gender neuter.
Type species. Allorapisma chuorum sp. nov.
Included species. The type species only.
Diagnosis. Forewing separable from those of all other ithonid genera except Principiala by proximal branches of Rs short, appear fused with MA; from Principiala by R1 space (between R1, Rs), much wider than subcostal space [contrary in Principiala] and costal gradate series of crossveins represented by few basal crossveins located near costal margin [extended to at least mid-point of costal space, located far from costal margin in Principiala].
Remarks. Some character states found in Allorapisma are of considerable interest.
The proximal branches of Rs are short and appear to fuse with MA. Within the family, this has been previously only found in the Cretaceous genus Principiala Makarkin & Menon (Makarkin & Menon 2007: Fig. 5A). This feature is also found in the vast majority of Chrysopidae (i.e., all Chrysopinae , Apochrysinae, and some Nothochrysinae , but absent in Mesozoic Lamiinae ), and possibly in the psychopsoid genus Hongosmylites Makarkin & Archibald, 2005. It appears clear that this condition is convergent among these three phylogenetically distant taxa, i.e., Ithonidae , Chrysopidae , and Psychopsoidea.
MA and MP are completely separated, with MA originating from R. This condition has been reported to occur very rarely within the order, and it is currently unclear if any of these cases may withstand close scrutiny. Makarkin and Menon (2007) considered M to be divided into MA and MP near the forewing base in Principiala incerta. Here we revise this interpretation. P. i n c e r t a is known from two specimens; in both, this region is somewhat obscured by the frenulum of the hind wing. Despite this, it can be confidently determined that in the forewing of the holotype of P. i n c e r t a, MA and MP indeed approach each other toward the wing base and it’s possible that they might have a common stem of M. In the paratype, however, MA appears to originate from R, i.e., MA is separated from MP ( Makarkin & Menon 2007: as seen in the photograph of basal portions of the wings in Makarkin & Menon 2007: Fig. 2 View FIGURE 2 B; the forewing was incompletely drawn in Makarkin & Menon 2007: Fig. 5A). Therefore, it should be considered that this character state may be variable in this species. Most Australian Ithonidae and some Eurasian Dilaridae (i.e., the majority of species of Dilar Rambur ) possess a superficially similar vein configuration in this area. In these taxa, one to three branches of what might appear to be MA also originate from R basal to the origin of Rs; here, we interpret these confidently as branches of Rs, as indicated of the location of the distal nygma, which in these families is always situated between the two most proximal branches of Rs (e.g., New 1991: Fig. 34.7A, for Ithonidae ; Monserrat 1988: Fig. 13–17, for Dilaridae ).
The basal fusion of MP and Cu is unique within the Neuroptera . If this is a consistent character state within the species and not an individual anomaly of the holotype (this part of the wing is missing from the paratype), it is unquestionably a significant autapomorphy.
The strong concavity of MA is unusual in hemerobiiform Neuroptera ; it is a condition more characteristic of psychopsoids (e.g., Kalligrammatidae , Osmylopsychopidae ) and myrmelentoids (e.g., Palaeoleontidae ) ( Menon & Makarkin 2008).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.