
Sivatherium garevskii, Geraads, 2009
|
publication ID |
https://doi.org/10.5252/g2009n4a893 |
|
persistent identifier |
https://treatment.plazi.org/id/03EF879F-FFB2-FFB3-C758-FE576C3BFDD7 |
|
treatment provided by |
Marcus |
|
scientific name |
Sivatherium garevskii |
| status |
sp. nov. |
Sivatherium garevskii n. sp.
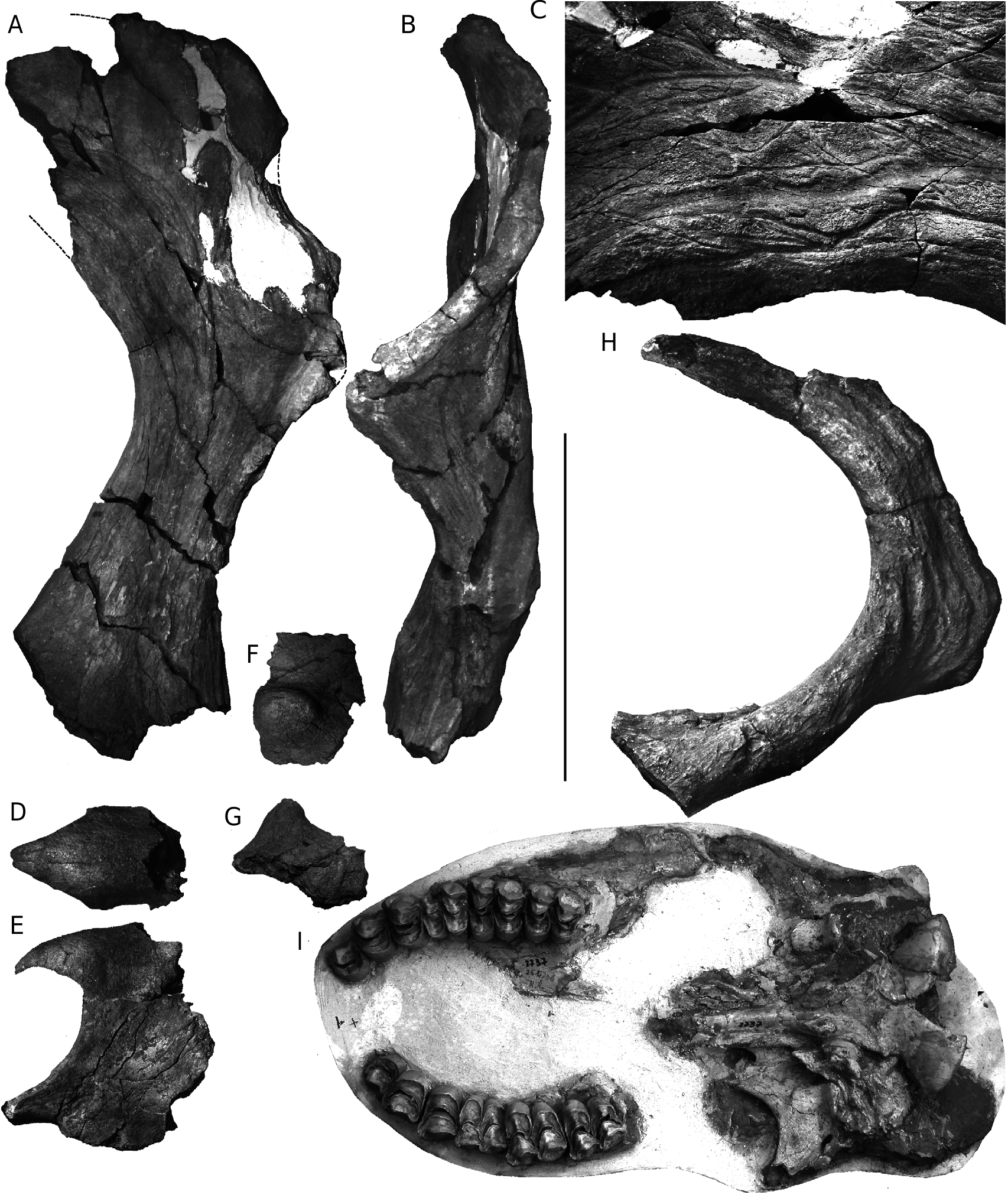
HOLOTYPE. — Fragments d’un crâne comprenant le nasal et une partie du maxillaire, deux dents supérieures, un appendice crânien antérieur et une corne postérieure gauche incomplète, sans numéro, conservés au MHNS ( Fig. 3 View FIG A-G).
ÉTYMOLOGIE. — En reconnaissance de la contribution majeure de Risto Garevski à la paléomammalogie macédonienne.
LOCALITÉ TYPE. — Stamer près de Delcevo, nord-est de la République de Macédoine. Le sédiment adhérant aux fossiles ne permet pas de choisir entre la Formation de Pancerevo, attribuée au Miocène supérieur, et celle de Solnje, attribuée au Pliocène ( Dumurdzanov et al. 2004); cependant, ce site a livré trois autres fossiles, une P3 référable à Propotamochoerus ? sp., genre mio-pliocène, et deux fragments de bois de cervidés n’appartenant sans doute pas à des genres modernes et, bien que très pauvre, un tel assemblage s’accorde mieux avec le Pliocène qu’avec le Miocène supérieur, où les cervidés sont rares et les bovidés dominants.
DIAGNOSE. — Sivatherium de taille moyenne d’après ses dents, mais à cornes principales gigantesques, courbées vers l’arrière, presque dépourvues de torsion, mais à bord antérieur formant un bourrelet ondulant, festonné de tubercules rugueux, à paumure atteignant sa largeur maximale vers la mi-longueur. Cornes antérieures petites, formées d’un simple tubercule. Prémolaires modérément développées. Sivatherium garevskii n. sp. se distingue des autres espèces du genre par par la petite taille de ses cornes antérieures, et par la taille beaucoup plus grande des cornes postérieures, dont la paumure s’élargit jusque vers la mi-longueur au lieu de diminuer de la base au sommet.
DESCRIPTION ET COMPARAISONS
Le genre Sivatherium comprend une forme asiatique, S. giganteum Falconer & Cautley, 1836 du Plio- Pléistocène des Siwaliks supérieurs (Indo-Pakistan), surtout connu par un crâne conservé au BMNH (M 15283), et deux formes africaines, S. maurusium (Pomel, 1893) (= S. olduvaiense Hopwood, 1934 )
du Pliocène moyen et supérieur et du Pléistocène, et S. hendeyi Harris, 1976 du Pliocène inférieur de Langebaanweg. La forme macédonienne se distingue bien de ces trois espèces, tant par son origine géographique que par ses caractères crâniens.
Le fragment de museau comporte sans doute la plus grande partie du nasal, basculé à 90° par rapport au prémaxillaire et à la partie rostrale du maxillaire, la suture entre ces os n’étant pas reconnaissable ( Fig. 3D, E View FIG ). Courts, un peu busqués et pointus en avant, les nasaux sont très semblables à ceux du crâne de S. giganteum (BM 15283); Colbert (1935: fig. 172) a représenté les nasaux de Bramatherium , Sivatheriinae plus ancien, sur le même modèle, mais ils sont en fait inconnus. Ceux du S. maurusium de Koobi Fora au Kenya ( Harris 1976b: pl. 15) sont également pointus, mais leur profil est presque rectiligne.
Le foramen infra-orbitaire était situé au-dessus de l’alvéole antérieur de P3, un peu plus postérieurement que chez S. giganteum et S. maurusium (Koobi Fora: Harris 1976b; Djibouti: Geraads 1985). Le fond de l’échancrure nasale se situe à l’aplomb de l’avant de P2, comme sur le specimen de Djibouti; il semble plus profond sur le specimen de Koobi Fora, et l’est assurément chez S. giganteum .
La corne postérieure ( Fig. 3 View FIG A-C) a été remontée à partir de nombreux fragments, mais leurs cassures très fraîches garantissent une reconstruction fidèle. Il manque néanmoins la partie distale, que j’évalue à un tiers au moins de la longueur totale. Son caractère le plus remarquable est sa très grande taille, dépassant nettement celle de tous les autres Giraffidae , y compris les plus grands Sivatherium ; chacune devait peser autour de 20 kg sur le vivant.
L’absence de torsion ne permet pas d’orienter la corne sans ambiguïté, mais à la base, la face la plus plane doit correspondre à la face médiale (dorsale), et il s’agit donc très probablement d’une corne gauche. La corne est donc recourbée vers l’arrière. Elle est fortement comprimée transversalement, le rapport entre son diamètre antéro-postérieur et le diamètre transversal (épaisseur) variant entre 1,4 au niveau de la constriction peu au-dessus de la base, et environ 4 au niveau où la corne est la plus étalée. Le bord postérieur (concave) est épais, de 10 cm au maximum près de la base, cette épaisseur dimi- nuant lentement vers le haut. La section transversale montrerait néanmoins, au dessus de la constriction où cette section est à peu près ovale, une diminution assez rapide de cette épaisseur vers le centre de la corne, de sorte que la plus grande partie est assez mince, l’épaisseur minimum étant d’environ 25 mm. Le bord antérieur est néanmoins un peu épaissi (environ 4 à 5 cm); il porte de plus des rugosités et nodosités irrégulières, qui contribuent à accentuer les variations importantes de l’étalement de la corne. Au-dessus de la constriction située peu au-dessus de la base, où la DAP n’est que de 140 mm, la corne s’étale en effet largement puisque cette DAP double, pour diminuer un peu, puis augmenter à nouveau (DAP maximale = 310 mm). Bizarrement, la DAP ne diminue donc pas plus ou moins régulièrement comme chez les Sivatherium africains, mais la paumure s’élargit au contraire vers le haut, comme chez le daim. Il est néanmoins presque certain que l’étalement diminuait dans la partie terminale manquante.
Si, en vue médio-dorsale ( Fig. 3A View FIG ), la corne semble à peu près plane, la vue antéro-latérale ( Fig. 3B View FIG ) montre que ce bord est affecté par une gracieuse ondulation qui le porte d’abord franchement en avant, puis le ramène dans le plan principal, puis le porte à nouveau un peu vers l’avant. Chez S. maurusium , au contraire, cette bordure décrit habituellement une courbe régulière, hétéronyme ( Fig. 3H View FIG ; Singer & Boné 1960: pl. 1a, c, d, e, pl. 3, pl. 29c, d; Harris 1974: fig. 4; 1991: fig. 4.25 Arambourg 1979: pl. 27, fig. 3, pl. 28, figs 6, 7; Geraads 1985: fig. 1). Dans de rares cas, la corne peut être dépourvue de torsion ( Singer & Boné 1960: pl. 1b, f; type C de Harris 1974), mais dans tous les cas la paumure se réduit vers le haut. Les cornes de S. hendeyi du Pliocène inférieur de Langebaanweg ( Harris 1976a) sont recourbées mais simples, presque dépourvues de tubérosités, et à peine aplaties. Malgré un âge peut-être voisin, elles sont donc très différentes de celles de S. garevskii .
Les cornes postérieures de S. giganteum sont mal connues. Falconer & Cautley (1836) n’en ont figuré que des fragments, le plus complet ( Murchison 1868: pl. 21, fig. 3, BMNH M 39525 View Materials ) étant une corne gauche, et supposée comme telle par ces auteurs et la plupart des auteurs ultérieurs, mais comme la figure est inversée, la reconstitution du crâne de S. giganteum était fausse, les cornes droite et gauche étant interverties, comme l’a noté Abel (1904). On y retrouve l’ondulation du bord antérieur, mais il existe en plus un fort «andouiller » postéro-externe (et non postéro-interne comme le montrent les figures de Falconer & Cautley 1836, et de Colbert 1935), totalement absent à Stamer et en Afrique. Un autre spécimen, figuré par Colbert (1935: fig. 171), comporte aussi des excroissances situées à l’opposé l’une de l’autre.
En Europe, le seul spécimen comparable à celui de Stamer est un fragment de corne de Turquie d’Europe, d’âge incertain mais sans doute pliocène, attribué par Abel (1904) à Sivatherium , et conservé au PIUW. Il ne s’agit que d’un tronçon, mais il correspond assez bien à celui de Stamer, malgré une courbure vers l’intérieur du bord postérieur. Les proximités géographique et, peut-être, chronologique, accroissent la probabilité qu’il s’agisse d’une même espèce.
L’absence d’érosion sur la plus grande partie de la corne de Stamer permet d’observer en détail les reliefs de sa surface ( Fig. 3C View FIG ). Aucune zone n’est lisse. Le bord antérieur est rugueux, mais tout le reste de la surface est couvert de sillons de largeur variable. Singer & Boné (1960: 495, 496) contestaient qu’il puisse s’agir d’empreintes vasculaires parce que leur diamètre ne diminue pas de la base au sommet, qu’elles ne sont pas liées aux foramens nourriciers, qu’elles ne sont pas plus fortes que sur les cornes antérieures qui sont bien plus petites, et que de tels vaisseaux seraient dangereusement exposés. Ces auteurs supposaient qu’ils auraient pu en fait servir d’ancrage à des attaches ligamentaires ou cartilagineuses d’un étui corné; cette hypothèse semble avoir été acceptée par Harris (1991: 117), mais elle est invraisemblable.Tout d’abord, la corne de Stamer montre bien qu’aucun des sillons présents vers l’extrémité supérieure n’atteint la taille des plus gros de ceux de la base; leur diamètre moyen diminue, même si d’apparentes anastomoses ne permettent pas de les suivre sur toute leur longueur. L’appendice crânien antérieur de Stamer ne porte que des sillons bien plus petits. Quant au risque d’exposer ces vaisseaux superficiels, les cervidés montrent qu’il est limité; il est probable que ces vaisseaux ne persistaient que pendant la phase de croissance de la corne, laissant ensuite, comme chez les cervidés, l’os à nu. La morphologie complexe de la corne et la présence de rugosités sur le bord antérieur excluent tout revêtement kératinisé.
Quelques fragments du toit crânien ont également été retrouvés, mais je ne suis pas parvenu à établir le contact entre la corne postérieure et la petite corne antérieure. Cette dernière forme un simple tubercule un peu allongé (antéro-postérieurement?), d’environ 85 × 75 mm pour 50 mm de hauteur (mesures approximatives, les limites de la corne étant très vagues).
La taille et la morphologie des appendices crâniens antérieurs diffèrent aussi de celles des autres espèces. Chez S. giganteum , ils méritent le nom de cornes, étant beaucoup plus longs, aplatis à la base mais coniques vers la pointe. Les appendices antérieurs de S. hendeyi , s’ils existent, sont inconnus. Harris (1974) avait attribué à cette espèce une corne longue et conique qui appartient sans doute en fait à un autre taxon, comme noté plus haut. Chez S. maurusium les appendices antérieurs sont aplatis, allongés, et peuvent être arrondis en vue latérale ( Geraads 1985) ou ensellés ( Singer & Boné 1960: pl. 29a, b; Harris 1991: fig. 4.20). On peut exclure l’éventualité que ces différences soient de nature ontogénique, car les dents de Stamer sont très usées, ou sexuelle, car les cornes postérieures du type de S. garevskii sont trop robustes pour être celles d’une femelle dont l’ornementation crânienne serait moins développée que chez les mâles; il se peut que les femelles aient été inermes, comme chez S. giganteum .
La dentition n’est représentée que par un fragment de molaire et une prémolaire, toutes deux très usées. Les dimensions de la prémolaire (L = 32, largeur = 42), probablement une P4, sont celles d’ Helladotherium ( Bohlin 1926) ou de S. maurusium ( Geraads 1996: tab. 1). La longueur de la rangée des prémolaires devait être inférieure à 100 mm; elle était donc plus courte que chez certains individus de cette dernière espèce, formes géantes récentes d’Afrique du Sud, ou au contraire formes les plus anciennes de Langebaanweg ( Harris 1976a) ou de Tunisie ( Arambourg 1979), et nettement plus courte aussi que chez S. giganteum . Ces dimensions modérées de la dentition contrastent avec le gigantisme des cornes.
DISCUSSION
La corne de S. garevskii de Stamer n’est que la seconde corne de Sivatheriinae découverte en Europe, plus d’un siècle après la première ( Abel 1904). Elle complique un peu la question de la nature réelle de l’ Helladotherium , connu depuis plus de 150 ans dans le Miocène supérieur, ou tout au moins le Turolien, de la région égéenne, à la suite des travaux de Gaudry & Lartet (1856) et de Gaudry (1862 -67) sur le matériel de Pikermi, mais dont le seul crâne adulte connu reste celui, inerme, décrit de ce site par ces auteurs. Puisque tous les autres Sivatheriinae , tant asiatiques qu’africains, possèdent des appendices crâniens bien développés, il est clair que le crâne de l’« Helladotherium » est celui d’une femelle d’un autre genre, dont la nature demeurait inconnue, les nombreuses fouilles menées dans le Miocène supérieur de la région égéenne n’ayant livré aucune corne (on peut soupçonner que si les restes d’appendices crâniens ont si rarement été signalés, c’est qu’ils sont sans doute bien peu reconnaissables sur le terrain, et que plus d’un ont été détruits au cours des fouilles). À la suite de la description d’un crâne juvénile du Miocène supérieur de Kavakdere en Anatolie, Geraads & Güleç (1999) ont conclu qu’il s’agissait sans doute de Bramatherium , genre jusque là limité aux péninsules indienne et arabique. Le crâne de Bramatherium , décrit et figuré par Bettington (1846), Colbert (1935) et Lewis (1939), est plus extraordinaire encore que celui de Sivatherium , puisqu’il présente deux paires de cornes, supra-occipitales et fronto-pariétales, ces dernières fusionnées à la base chez l’adulte, et dirigées presque verticalement. On peut néanmoins fort bien concevoir que le crâne de la femelle en ait été dépourvu, et de ce fait, attribué à un genre différent. Bramatherium – Helladotherium serait donc l’un des rares genres de Ruminants (avec par exemple les Bovidae Tragoportax Pilgrim, 1937 et Nisidorcas Bouvrain, 1979 ) communs à la région balkanique et au sous-continent indien au Miocène supérieur.
La confirmation, à la suite d’Abel, de la présence d’un autre sivatheriiné en Europe orientale, S. garevskii , soulève maintenant la possibilité que Helladotherium soit en fait plus voisin de Sivatherium . Bohlin (1926) avait montré que la femelle de S. giganteum est un autre «genre » asiatique, Indratherium , distinct d’ Helladotherium , mais ceci ne permet pas d’exclure que ce dernier soit la femelle de S. garevskii . Cette hypothèse me semble néanmoins peu probable, parce que le giraffidé de Stamer est sans doute plus récent que Helladotherium , et surtout parce que Bramatherium est un candidat bien plus satisfaisant à ce poste ( Geraads & Güleç 1999).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.