
Chaeropus ecaudatus ecaudatus Ogilby 1838
|
publication ID |
https://doi.org/10.11646/zootaxa.4566.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4D8BAEF5-AED6-4973-897C-96C5945E3710 |
|
DOI |
https://doi.org/10.5281/zenodo.5584110 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE2B15-D94F-FFAD-4C81-B5FBA41B24A2 |
|
treatment provided by |
Plazi |
|
scientific name |
Chaeropus ecaudatus ecaudatus Ogilby 1838 |
| status |
stat. nov. |
Chaeropus ecaudatus ecaudatus Ogilby 1838 View in CoL new status
( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Holotype. AM PA422 (previously AM Bennett’s Cat. 35), juvenile female skull, collected by Sir Thomas Mitchell on the Murray River , New South Wales.
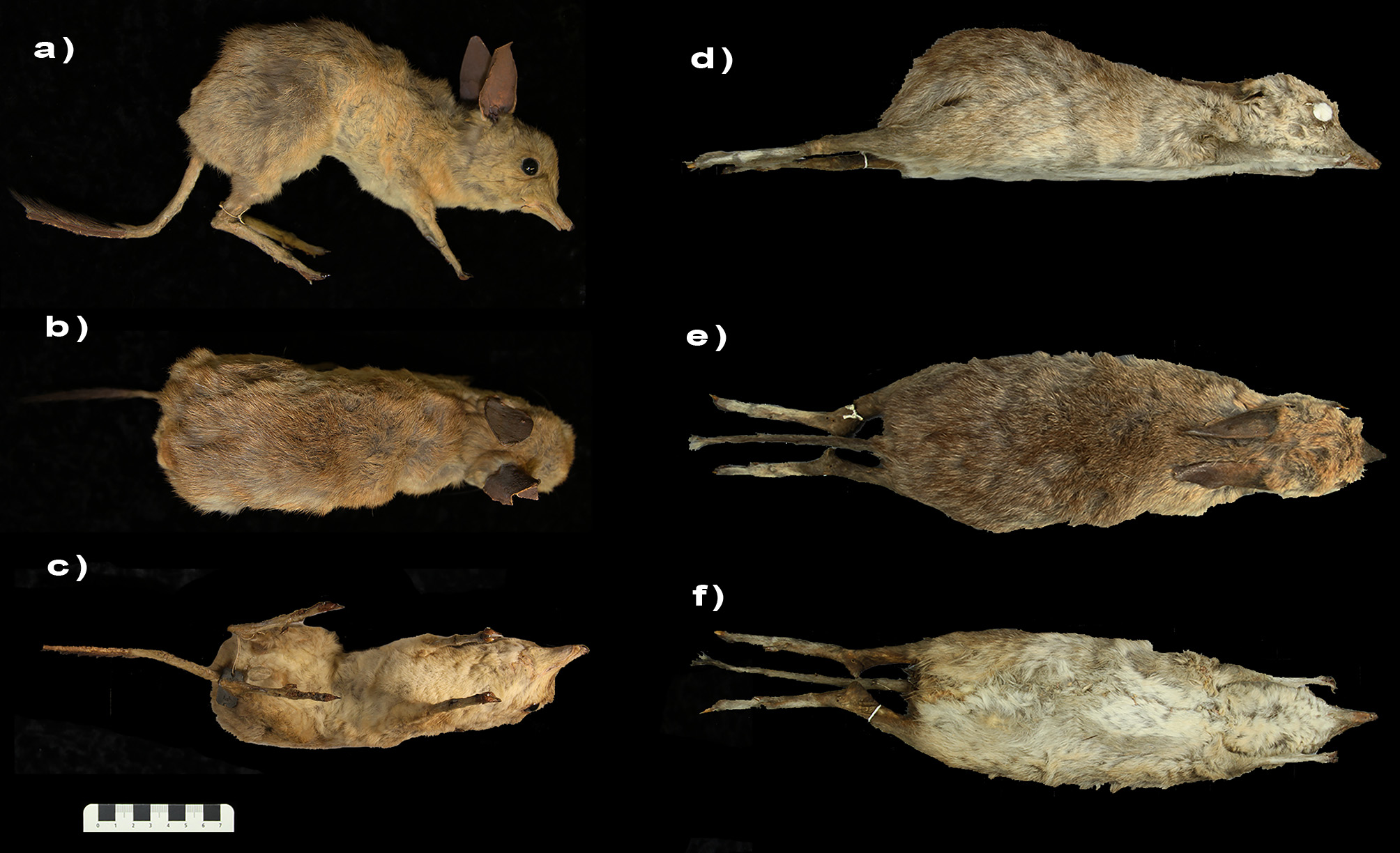
Referred specimens (modern). AM PA421 , adult male, taxidermy ; NHMUK ZD 1842.2 About NHMUK .1.33, holotype of C. castanotis , study skin with associated partial skull and dentaries ; NHMUK ZD 1847.8 About NHMUK .14.12, female, study skin with associated skull and dentaries ( NHMUK ZD 1848.1.27.41) ; NHMUK ZD 1847.8 About NHMUK .14.13, study skin with associated skull ( NHMUK ZD 1848.1.27.40) ; NHMUK ZD 1847.8 About NHMUK .14.46, postcranial material ; NMV C2900 About NMV , skull with associated dentaries ; NMV C5886 About NMV , juvenile, taxidermy ; NMV C5887 About NMV , adult male, taxidermy ; NMV C5889 About NMV , adult female, taxidermy ; NMV C27998, adult female, taxidermy ; NMV C36365, taxidermy .
Referred specimens (fossil). From late Pleistocene of Lake Menindee, New South Wales, Site 3, V67186 View Materials ; UCMP 52667 View Materials , two left dentaries, one with m2, other with m2–4 ; UCMP 52668 View Materials , right dentary with m2–4; from middle to late Pleistocene Mt Etna , Queensland, QML1312 ; QMF51637, right maxilla with M2– 4; QMF51638, right M2; QMF51639, right M3; QMF51640-51641 (specimens from the Queensland Museum were not directly examined but identified from photos in Hocknull 2005) .
Emended diagnosis. Chaeropus ecaudatus ecaudatus differs from C. e. occidentalis in having beige fur ventrally; shorter ears ( 50–56 mm versus 60mm in C. e. occidentalis ); shorter medial accessory palatal fenestrae; thick bridge between the maxillopalatine fenestrae and maxillary fenestrae; and larger upper and lower premolars and molars (see result section). C. e. ecaudatus differs from C. yirratji sp. nov. in having shorter hindfeet and tail; having maxillary fenestrae; having distinct medial accessory palatal fenestrae; having the paracone connecting to both StA and StB on M1 (in unworn teeth); having a reduced metaconule on M1–3, making the molars more triangular in shape (most noticeable on M3); carotid foramen opens ventrally (versus ventrolaterally in C. yirratji sp. nov.); StD1 absent on M1; metacone of M4 is distinctively larger; and the paracristid on m1 is continuous.
Remarks. The holotype, AM Bennett’s Cat. 35, has been noted as missing from the Australian Museum collections for quite some time (e.g. Mahoney & Ride 1988; Parnaby et al. 2017). Parnaby et al. 2017, who discuss the history of the missing specimen, mention that Krefft (1864) lists two specimens from the Australian Museum, an adult male from “Gall Gall Creek”, which originally came from Museum Victoria, which matches specimen AM PA421, and a skull from lower Murray, Sir Thomas Mitchell’s original specimen, which is most likely AM PA422. Krefft (1870) also mentions finding the skull “ten years ago” and thought the skin may have been disposed of. Even though Troughton (1932) believed the holotype missing from the collection, he may have been referring to the skin. AM PA421 is an adult male collected on the Blandowski expedition ( Wakefield 1966), and as is the case for all of the taxidermy prepared at Museum Victoria (NMV C5887, C5889 and C27998), the skulls were not kept ( Krefft 1864). As a result, AM PA422 cannot be associated with AM PA421, and is most likely Sir Thomas Mitchell’s original specimen as Krefft (1864) suggested.
Specimens from Mt Etna are tentatively placed in this subspecies due to the presence of the maxillary fenestrae below M1–2, and closer proximity to the type locality than the other subspecies. However, the M2 has a more developed metaconule and the postmetaconulecrista extends further posteriorly on the tooth relative to other specimens of Chaeropus ecaudatus ecaudatus and in this respect is similar to C. yirratji sp. nov.
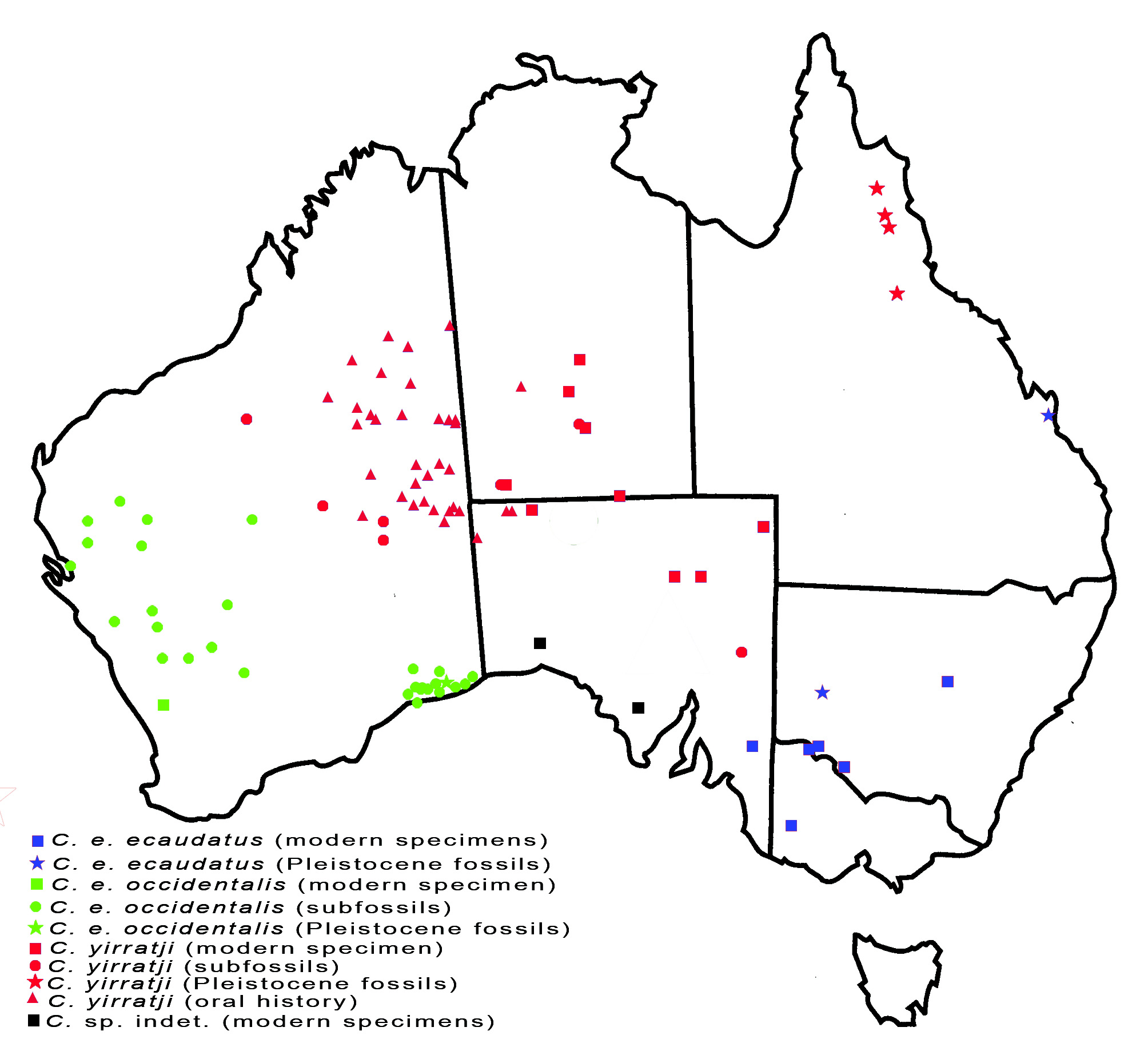
Distribution. Arid zone of western New South Wales, northern Victoria and south-eastern South Australia ( Fig. 1 View FIGURE 1 ).
Description. The following description is based on the holotype (AM PA422) and the referred specimen (AM PA421), unless otherwise specified. The holotype is a skull of a juvenile female, nearly adult, with dP3/dp3 still present, M4 not erupted, but m4 erupted. The referred specimen is a taxidermied adult male. Pelage is chestnut dorsally, gradually becoming more ochre laterally and beige ventrally. The pelage is similar in specimens from Museum Victoria (prefix NMV), but other specimens from the NHM (e.g. NHMUK ZD 1842.2.1.33, 1847.8.14.12, 1847.8.14.13) are often much darker in colour (perhaps less faded due to the way specimens were treated or stored), with a darker chestnut dorsally, light brown or grey laterally, and light brown to grey ventrally. Head is rounded, with short pointed snout. Ears are c. 50 mm long (about as high as the height of the skull). Forelimbs are shorter than the hindlimbs. On the forefeet, digit II and III are well developed with an ovoid foot pad. Digit IV is heavily reduced, located more posterior to digit II and III on the feet, and clawless. Digits I and V are not present. On the hindfeet, digit IV is well-developed with a large foot pad. Digit I is not present, and the syndactylous digits II and III, and digit V are heavily reduced and located more posterior to digit II and III on the feet. Tail is shorter than the body ( 190 mm in length), and crested.
The holotype skull is 48 mm long and 25 mm wide. In dorsal view, the nasals increase in width posteriorly, and the posterior edge of the nasal bones is located just anterior to the anterior margin of the orbits. The nasals do not extend anteriorly much further than the premaxillary-nasal suture. The maxilla contributes to a large portion of the ventral margin of the orbit. The parietals are fused at their dorsal margin as in all peramelemorphians. The anterodorsal margin of the orbits are formed by the lacrimals and are heavily crested (seen in both dorsal and lateral view), with a single lacrimal foramen sandwiched between the lacrimal crest and the lacrimal-maxillary suture. In lateral view, the auditory bullae are large ( 6.3 mm in height), rounded anteriorly but more pointed ventrally. The infraorbital foramen is located just above the anterior of M1, with an ovoid incisivomaxillary canal just anterior to it, which pierces the maxilla anteromedially. The anterior margin of the premaxilla, bordering the nasal cavity is straight in lateral view. The zygomatic arch thins to a minimum midway along its length, but is much thicker in dorsoventral height at its anterior and posterior ends. The antorbital fossa is wide and deep, extending anteriorly from a point just posterior to the infraorbital foramen to just posterodorsal to the posterior edge of M3 (on the zygomatic arch). A distinct ventral crest, continuous with the ventral edge of the zygomatic arch, separates the antorbital fossa from the tooth row. In ventral view, the palate is marked by having a number of vacuities. The incisive foramina taper anteriorly and are widest near their posterior edge. They extend from the anterior of I3 to the anterior of C1. The premaxilla borders the incisive foramina, except at their medial and posterior most margins, which are bordered by the maxilla. The accessory palatal fenestrae are large and represent two pairs of fenestrae, bordered entirely by the maxilla. The larger lateral pairs extend from the anterior of P1 to the middle of P2. The medial pair is smaller, about half the size of the lateral pair, and level with the posterior half of the lateral pair. In NMV C2900 and NHMUK ZD 1848.1.27.41, two additional small fenestrae (anterior medial accessory palatal fenestrae) are present anterior to the medial fenestra, but only one is present in NHM 1848.1.27.40. The maxillopalatine fenestrae are much larger than the accessory palatal fenestrae but are heavily damaged in the holotype, so only the anterior and lateral margins are visible. The anterior margins are concave posteriorly, and level with the dP3 (with P 3 in adults). The lateral margins are relatively straight for most of their length, ending roughly level with the posterior of M1. Maxillary fenestrae are present lateral to the maxillopalatine fenestrae and medial to M1–2. They are oval in shape and extend from the metaconule of M1 to just anterior to the metaconule of M2. The palatine fenestrae are not preserved in the holotype. They are only completely preserved in NMV C2900, but they seem to be variable in shape and size. Additional small vacuities are present throughout the palate, but these are likely to be the result of the fact that the palate was still developing in the holotype at the time of death, and these are not present in any of the adult specimens. There is a clear gap between the presphenoid and the basisphenoid, and the palatine does not form a wall at the presphenoid-basisphenoid suture (it completely encloses that suture in Macrotis , Isoodon and Perameles ), but instead opens up as a large transverse foramen. The carotid foramen is oval in dorsal view, opening ventrally, lacking a distinct ventral wall (opens laterally in all other peramelemorphians). A distinct groove occurs in the basisphenoid directly posterolateral to the carotid foramen, likely for the attachment of nerve and blood vessels. The transverse foramen is directly lateral to the carotid foramen and more circular in shape. The primary foramen ovale is oval in shape and located medial to the anteromedial ‘corner’ of the auditory bullae and is completely enclosed by the alisphenoid. The secondary foramen ovale is slightly anterolateral to the primary foramen ovale, and opens anteriorly. An additional smaller foramen is present just lateral to the secondary foramen ovale (also present in Isoodon , but absent in Perameles ). The promontorium of the petrosal is marked by a high crest, oriented anteromedial-posterolateral. The dentary has a unique feature (not present in any other marsupial) on the base of the ascending ramus (in lateral view), a triangular shaped process, which would have been used for additional attachment for the masseter muscle.
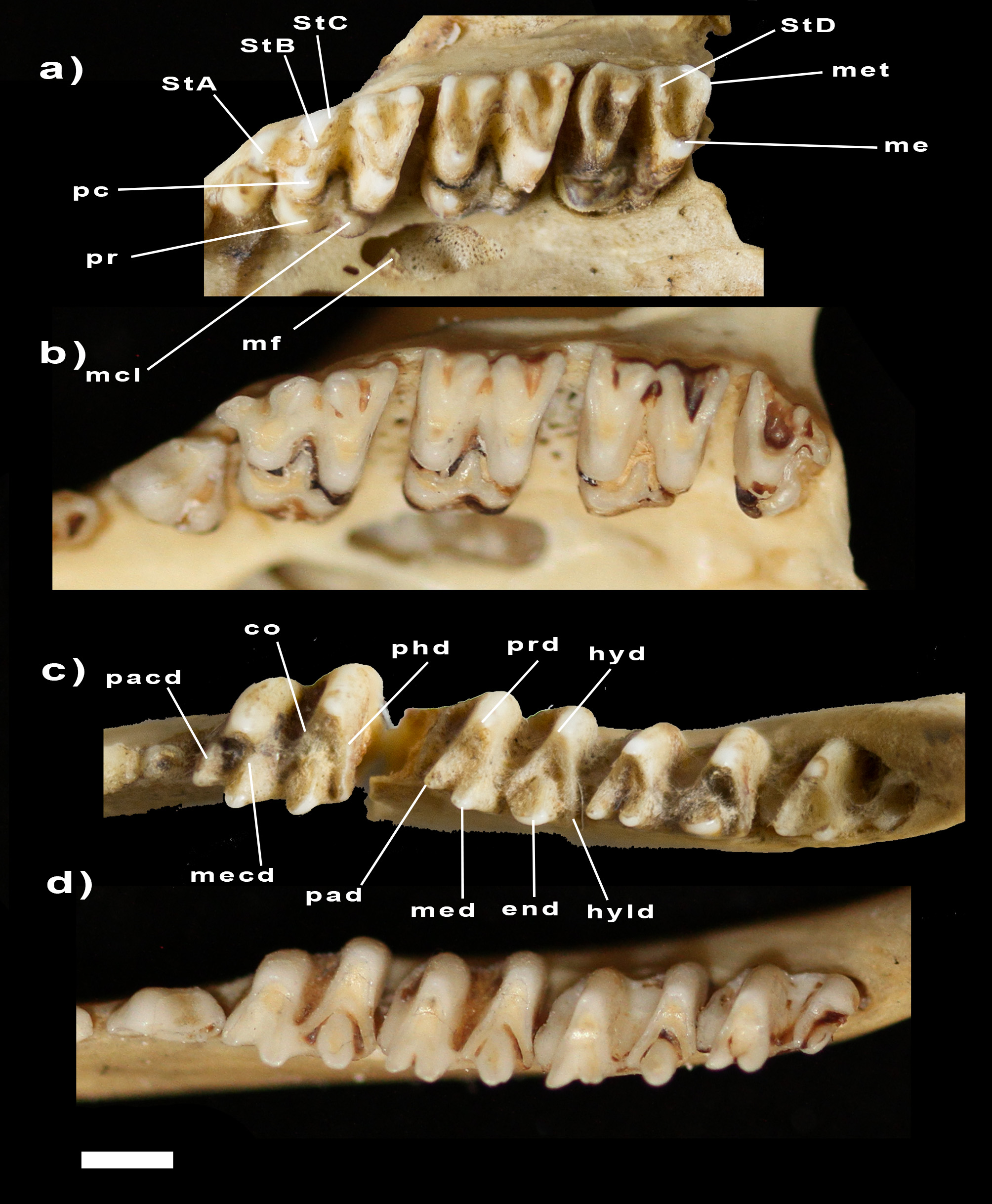
The five upper incisors are rhomboidal in shape, increasing in size from I1 to I3 then decreasing in size from I3 to I5. A short diastema (shorter than the length of C1) is present between I5 and C1. The C1 is premolariform, double rooted, with distinct anterior and posterior cusps. C1 is smaller than P1. A short diastema separates C1 from P1, and a shorter diastema separates P1 and P2. P1 and P2 are similar in shape, with a large central cusp, a small anterior cusp, and a larger posterior cusp connected to the large central cusp by a blade. P2 is larger (longer, wider and taller) than P1. The dP3 is a small rounded tooth, with a central cusp and two blades descending the crown, one laterally, and the other posterolaterally. The P3 is preserved in adult specimens (e.g., NMV C2900). It is longer and much wider than P2, with a large buccal cusp and a posterolateral shelf with a large cusp. The M1 is the longest molar on the tooth row; it is less wide and less high crowned than the posterior molars. StA is very large and connects via the preparacrista posteromedial surface of the paracone. The paracone is also connected to StB, an ovoid crested cusp. A small StC is visible posterior to StB in buccal view. The postparacrista is oblique and ends posterolingual to StB. The metacone is posterior to the paracone and connects to the base of StD via the premetacrista and to the tip of the metastyle via the postmetacrista. A remnant of StE is present posterior to the metastyle as a short crest. No crest connects StD to the metastyle. StD1 is absent. The protocone is lateral to the paracone, and a very low talon. The preprotocrista ends just anterior to the base of the paracone. The postprotocrista is long and extends into the valley between the paracone and metacone as a tall crest. The metaconule is smaller than the protocone and directly posterior to it. The premetaconulecrista is well-developed and distinct to the postprotocrista. The postmetaconulecrista is short and ends at the lingual flank of the metacone. The M2 and M3 are similar in shape and only differ from the M 1 in the morphology of the parastylar shelf. StA is small and more posteriorly positioned, lengthening the preparacrista and changing its direction (from that of the M1) to perpendicular to the tooth row. The postparacrista connects to StB. The metaconule is smaller in M2 than in M1, and even smaller in M3, making the teeth more triangular in shape. The M4 isn’t preserved in the holotype and was unlikely to have been fully erupted. Adult specimens conserve the M4. The parastylar shelf is longer and narrower than on M3. The postparacrista does not connect to StB, but instead curves posteromedially at the base of StB and joins to an extremely reduced metacone. The protocone is smaller and the talon is reduced, ending just below StB.
The three lower incisors increase in size posteriorly, with the i3 being bilobed as in all peramelemorphians. The crowns are flat and bladed. The c1 is directly posterior to i3, and premolariform, with large anterior and posterior cuspids on either side of the central cusp, but the tooth is shorter than the p1. Very short diastemata are present between the c1, p1 and p2. The p1 and p2 are similar to the c1, increasing in size posteriorly. The anterior cuspid on p1 and p2 is larger than the posterior cuspid, and with its own blade. The dp3 is a very small tooth, with a small anterior cusp, and a concave posterior blade running along the edge of the tooth. In adult specimens, the p3 is present and is shorter than the p2, but wider. The m1 is longer than it is wide, with a higher crown on the buccal side compared to the lingual side of the tooth. The trigonid is wider than it is long, with a small paraconid connected to the protoconid by a continuous paracristid. The protoconid connects to a large metaconid via the metacristid. The hypoconid is positioned more buccal than the protoconid. Cristid obliqua ends directly posterior to the metaconid. The posthypocristid is straight and ends at the very small hypoconulid. The entoconid is triangular in shape, with a preentocristid descending anteriorly, and a postentocristid descending buccally. The m2 and m3 are similar to one another and only differ from the m 1 in having a wide anterior cingulum, a wider trigonid and a larger entoconid. The m4 is similar to the m2 and m3 except the trigonid is longer and less wide, and the talonid is heavily reduced in size. In NMV C2900, the posthypocristid of the m4 is further reduced and ends posterobuccal to the entoconid.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |