
EULIPOTYPHLA
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00614.x |
|
persistent identifier |
https://treatment.plazi.org/id/03EAAD4B-FF84-135E-FECE-FB0FFE167229 |
|
treatment provided by |
Valdenar |
|
scientific name |
EULIPOTYPHLA |
| status |
|
Soricomorpha s.s. (e.g. not including tenrecs, etc.) There are some basic similarities between shrews and apatemyids in the form of the I1. Also, material of Heterohyus was published by Filhol (1890) as Neosorex , a supposed fossil shrew, highlighting the presence of some (limited) dental similarities between these groups. Soricomorphs possess several derived features of the skull that are missing in apatemyids, however, including a persistent, large piriform fenestra, an incomplete zygomatic arch, and a doubled mandibular condyle. Perhaps more importantly, shrews are lacking one of the most distinctive features of the basicranium of L. kayi : the basisphenoid tympanic process. Optimization of this feature is equivocal for the group that includes shrews, hedgehogs, and moles (Asher, 2005); it may be primitive for this group, suggesting that its absence in modern shrews may not be a bar to its presence in an ancient fossil relative. However, there are no features of the skull that offer clear support for an apatemyid– soricomorph relationship.
Of all of the taxa to which L. kayi was compared, the greatest phenetic similarities are with modern erinaceid insectivores, and in particular with Hemiechinus . Similarities between these forms include: (1) a horizontal ectotympanic; (2) caudal and rostral processes of the petrosal that are small but distinct; (3) a small alisphenoid tympanic process, through (or past) which the ramus inferior of the stapedial artery passes; (4) an un-enclosed facial nerve; (5) no complete ossified bulla; (6) a limited contribution of the petrosal to the tympanic roof; (7) grooves on the promontorium for the branches of an unreduced internal carotid artery, proceeding from a posteromedial entrance to the middle ear; (8) a substantial contribution from the sphenoid to the tympanic roof; (9) a basisphenoid tympanic process; and (10) a small or absent piriform fenestra. Some of these features (e.g. 4, 5, 6, and 7) are likely to be primitive (based on their presence in primitive eutherians); for others (e.g. 1 and 10) the polarity is in debate ( Symonds, 2005). One of these traits (9) has often been highlighted as a potential synapomorphy of either Lipotyphla , or of a more restricted grouping within that cluster of forms (Asher, 2005; Symonds, 2005); however, the distribution of this character makes its significance uncertain (Asher, 2005).
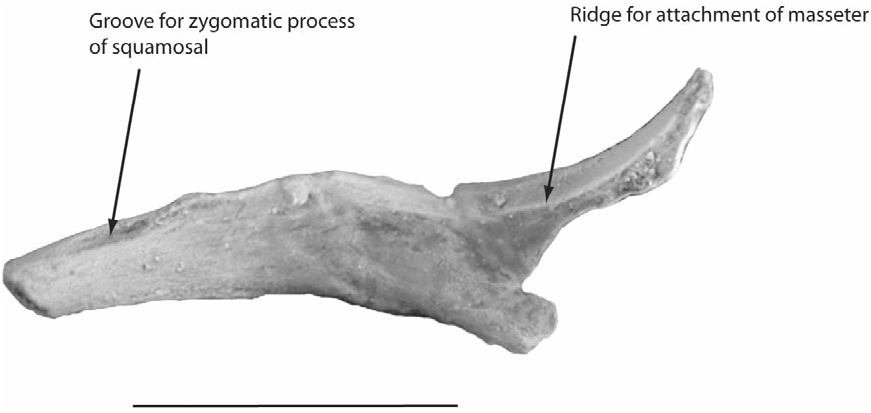
There are additionally some fairly significant differences between erinaceomorphs and apatemyids. A potential lipotyphlan synapomorphy is the reduction of the jugal ( Symonds, 2005). Erinaeomorphs (talpids and erinaceids) do retain a complete zygomatic arch (unlike soricomorphs); however, the jugal component of the arch is quite small. This is not true of apatemyids, in which the jugal is a very sturdy element (e.g. see Fig. 15 View Figure 15 ), and the zygomatic arch is quite robust. However, there may be a functional explanation for this characteristic. As discussed above, the zygomatic arch probably had an important role to play in resisting the bending forces in the face resulting from using the incisors to bore into wood. As such, this difference may have more to do with function than phylogeny.
A feature reconstructed as primitive for Lipotyphla is an expansion of the maxilla into the orbital mosaic (Asher, 2005), with the expansion of this bone being particularly marked in erinaceomorphs. Labidolemur lacks this trait ( Fig. 16 View Figure 16 ): the maxilla does not extend to the level of the dorsal edge of the infraorbital foramen. Another feature of some eulipotyphlans that has been considered significant (e.g. McDowell, 1958; see discussion in Boyer & Georgi, 2007) is the absence of a ‘true’ postglenoid process, although this trait is quite variable in insectivores, and is present in some fossil members of the group ( Butler, 1988). In any case, apatemyids do have a lip-like postglenoid process, which is located caudolateral to the postglenoid foramen, rather than medial to it as in some erinaceomorphs. In sum, with the exception of the basisphenoid tympanic process, apatemyids lack most of the features that have been considered as potential synapomorphies for Lipotyphla or Eulipotyphla .
There are other differences between apatemyids and living erinaceomorphs, including the breadth of the ectotympanic (broader in erinaceomorphs), the degree of development of the basisphenoid tympanic process (more laterally extensive in erinaceomorphs, although this may be influenced by breakage in L. kayi ), and the size of the premaxilla (much smaller in erinaceomorphs). Also, L. kayi is actually more similar to living erinaceomorphs than it is to some fossils that have been suggested to be related to that group. Brachyerix , for example, is very different from both modern erinaceomorphs and L. kayi in having a much more heavily ossified auditory region, with canals for the internal carotid artery, and an even more expansive basisphenoid tympanic process, forming an ossified auditory bulla ( Rich & Rich, 1971). Diacodon actually lacks the basisphenoid process ( MacPhee et al., 1988), further complicating our picture of the evolutionary history of that trait.
In sum, comparisons with a diverse array of living and extinct eutherian groups, reveals no obvious solution to the problem of apatemyid relationships. While similarities to erinaceomorphs are intriguing, their polarity needs to be assessed in the context of a phylogenetic analysis.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |