
Halticus, HAHN, 1832
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00770.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E8878D-FFC3-FFD3-5F14-FCABB066FC18 |
|
treatment provided by |
Marcus |
|
scientific name |
Halticus |
| status |
|
HALTICUS HAHN View in CoL View at ENA ( FIGS 3 View Figure 3 , 30–32 View Figure 30 View Figure 31 View Figure 32 )
Astemma Latreille, 1829: 199 (nom. nud.); junior homonym of Astemma Lepeletier & Serville, 1825 [ Heteroptera : Pyrrhocoridae View in CoL ]; type species: Acanthia pallicornis Fabricius, 1794 = Cicada aptera Linnaeus, 1758 by monotypy; Westwood, 1840: 121 (description); Amyot & Serville, 1843: 284 (description)
Halticus Hahn, 1833: 113 View in CoL (gen. nov.; type species: Acanthia pallicornis Fabricius, 1794 = Cicada aptera Linnaeus, 1758 by monotypy); Burmeister, 1835: 277 (descr.; Spinola, 1850: 43 (key); Fieber, 1858: 312 (key); Fieber, 1861: 69, 281 (key; descr.); Reuter, 1875a: 23 (key); Reuter, 1875b: 86, 89 (key; descr.); Saunders, 1875: 287 (key); Atkinson, 1890: 117 (cat.); Reuter, 1891: 17, 157 (descr.; key); Saunders, 1892: 267 (key; descr.); Distant, 1904: 479 (descr.); Kirkaldy, 1906: 130 (cat.); Hueber, 1906: 2, 42 (key; descr.); Oshanin, 1910: 802 (cat.); Reuter, 1910: 147 (cat.); Poppius, 1914: 84 (key; descr.); Van Duzee, 1916: 211, 374 (cat.); Knight, 1923: 498 (key); Blatchley, 1926: 800 (eastern US species key); Stichel, 1933: 235 (Palaearctic key); Hedicke, 1935 (Palaearctic key); Knight, 1941: 17, 75, 77 (Illinois key; descr.); Hsiao, 1942: 253 (Chinese key); China, 1943: 268 (nomenclature); Zimmerman, 1948: 199 (Hawaiian key); Slater, 1950: 45 (female genitalia); Kiritshenko, 1951: 127 (USSR key); Wagner, 1952: 95, 96 (Palaearctic key; descr.); Carvalho & Leston, 1952: 245 (British key); Carvalho, 1952: 74 (list); Carvalho, 1955: 65 (key); Carvalho, 1956: 51 (key Micronesian spp.); Carvalho, 1958: 12 (cat.); Southwood & Leston, 1959: 244 (British key); Kelton, 1959: 33, Fig. 81 (male genitalia); Wagner, 1961: 48 (diag.; Palaearctic key); Kerzhner, 1964a: 968 (diag.; USSR key); Maldonado, 1969: 57 (diag.; US key); Alayo, 1974: 15 (diag.); Schuh, 1974: 29 (discussion); Wagner, 1973: 6 (descr.; Palaearctic key); Kelton, 1980: 194 (diag.); Henry, 1983: 610 (Western Hemisphere key); Linnavuori, 1994: 5 (African key); Cassis & Gross, 1995: 186 (synonymy; Australian cat.); Schuh, 1995: 53 (synonymy; world cat.).
Eurycephala Laporte, 1832: 36 (gen. nov.; Lygaeus luteicollis Panzer, 1805 by subsequent designation Kirkaldy, 1906: 170); Brullé, 1836: 140 (descr.); Spinola, 1840: 191 (descr.); Blanchard, 1840: 140 (descr.); Reuter, 1891: 17 (synonymy); Carvalho, 1958: 12 (cat.)
Halticocoris Douglas & Scott, 1865: 478 (unnecessary nom. nov. for Halticus Hahn View in CoL nec Haltica Illiger [ Coleoptera View in CoL ]); Carvalho, 1958: 13 (cat.);
Halticiellus Villiers, 1952: 186 (gen. nov.; type species: Halticus tibialis Reuter, 1891 View in CoL by monotypy); Carvalho, 1958: 12 (synonymy)
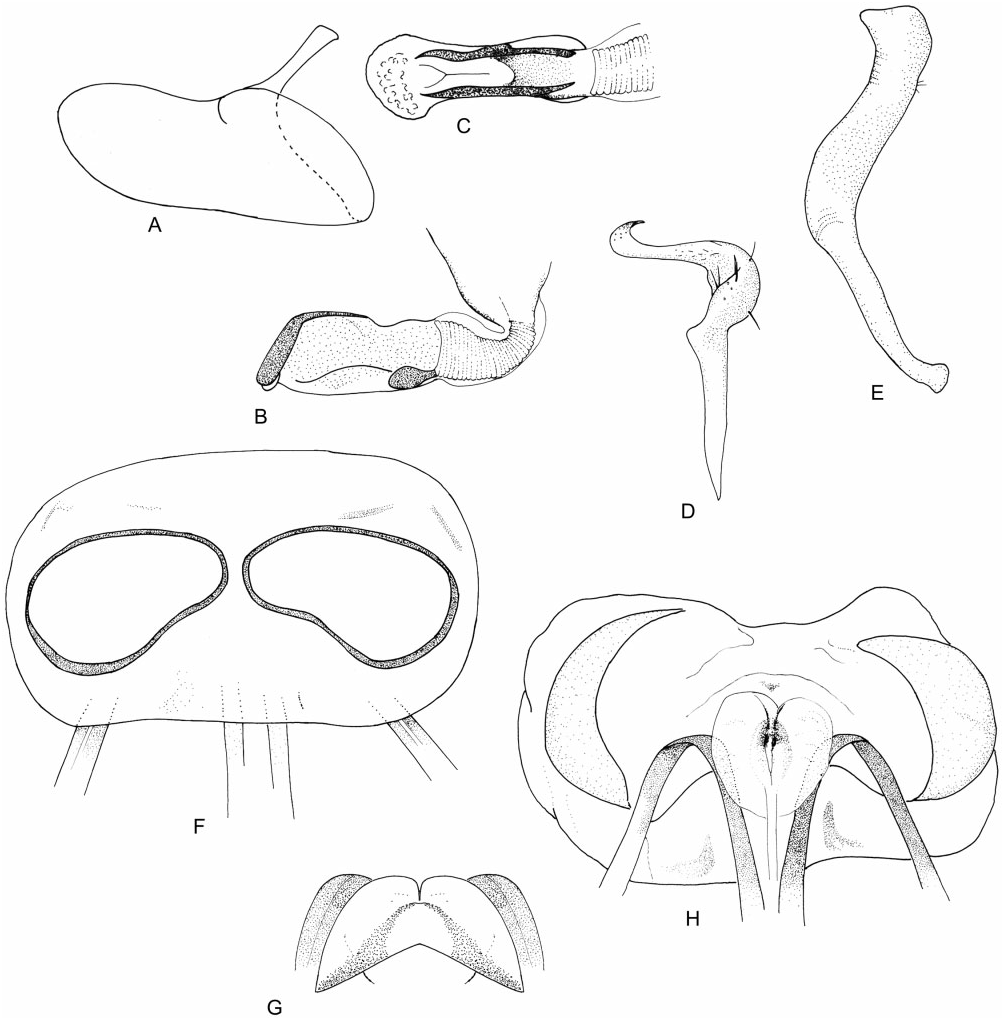
Diagnosis: Halticus is characterized by a combination of the following: length of first labial segment less than gena height; elongate, flattened right paramere; left paramere with twisted apophysis; scoop-shaped secondary gonopore and vestigial endosoma lacking spicules; large, thin sclerotized rings.
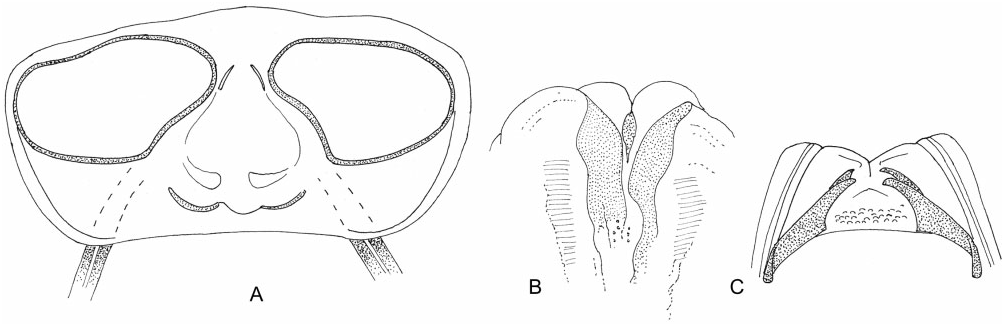
Redescription: Usually small and robust, macropterous or coleopteroid. Coloration ( Fig. 3 View Figure 3 ): almost all species black, sometimes with paler yellow, orange, red, or brown colouring on head, antennae, and legs, rarely uniformly brown. Surface and vestiture ( Figs 3 View Figure 3 , 30A–E, G, H View Figure 30 ): variable, generally glossy, sometimes wholly impunctate, in others pronotum is shallowly punctate; frons and vertex with faint creases radiating from centreline. Body generally with two forms of setae – thin and simple and white, flat, and scale-like; antennae and legs with short semi-erect spines and spine-like setae. Structure: head ( Figs 3 View Figure 3 , 30A, B, D View Figure 30 ): transverse and short, ranging from broader than tall to taller than broad; vertex with transverse sulcus, posterior margin carinate, concave, generally broader than anterior of pronotum; head height variable, usually> two times eye height; eyes sometimes wrapping around pronotum, sometimes touching; eyes tall and short; clypeus narrow, dorsal margin weakly sulcate; mandibular plate depressed; maxillary plate raised and carinate. Labium ( Fig. 30D View Figure 30 ): length variable, LI greatly swollen. Antennae ( Figs 3 View Figure 3 , 30A–D View Figure 30 ): insertion typically in line with or slightly below lower margin of eye; long, thin, and whip-like; AI slightly thicker than other segments, about as long as eye height. Thorax ( Figs 3 View Figure 3 , 30A, D–F View Figure 30 ): pronotum trapezoidal (macropterous) to subrectangular (coleopterous), weakly declivent, laterally sloping, collar thin and flat to absent; callosite region ill-defined to obsolete, posterior margin thinly carinate, straight to weakly medially cleft; metathoracic spiracle prominent, narrowly bordered with evaporative bodies; MTG external efferent system swollen, occupying lower half of metepimeron, ostiole opens laterally, peritreme rounded, orientated posteriorly, surrounded by evaporative bodies. Hemelytra ( Fig. 3 View Figure 3 ): macropterous: costal margin weakly curved, broadest above cuneal fracture; clavus longitudinally folded, with lateral side steeply sloping; corium laterally sloping; membrane and cuneus deflected. Coleopterous: broadly rounded, laterally and posteriorly sloping; claval suture and medial fracture faint, cuneal fracture very short. Legs: long, metafemur greatly swollen; pretarsi with fleshy pulvilli. Male genitalia ( Figs 30H View Figure 30 , 31A–E View Figure 31 ): pygophore conical, posterior margin sinuate, with broad deep concavity below left paramere; phallotheca simple and elongate-oval; left paramere apically twisted with sinuate apophysis; right paramere longer than left, extends out of pygophore, broad, flat, sometimes angled laterally at midpoint, apex rounded with a small apicolateral apophysis; ductus seminis attenuate, with flexible ribbing; secondary gonopore partly sclerotized, in form of elongate scoop, nearly filling phallotheca; endosoma extremely reduced, without spicules. Female genitalia ( Figs 31F–H View Figure 31 , 32A– C View Figure 32 ): dorsal labiate plate mostly membranous; sclerotized rings large, transverse, medially subcontiguous, sclerotized margins thin, lateral margins weakly upturned; VLP weakly sclerotized on posterior margin, laterally sometimes with paired, scimitarshaped sclerotizations, curving laterally and reaching up to dorsal labiate plate; posterior wall of bursa copulatrix variable, for the most part membranous, often with narrow paired sclerotized bands or rods convergent anteriorly; vestibulum symmetrical, with medial margins of first gonapophyses swollen outwards and sclerotized, vestibular opening rounded and strongly sclerotized.
Diversity and distribution: Halticus has 16 species and a mainly Holarctic distribution, with most species confined to the Palaearctic. A single species, Halticus tibialis , is found in Africa, South-East Asia, and the south-west Pacific.
Included species: Halticus apterus ( Linnaeus, 1758) * Palaearctic
Halticus asperulus Horváth, 1898 View in CoL Armenia
Halticus beganus Linnavuori, 1984 View in CoL Iraq
Halticus darbandikhaus Linnavuori, 1984 View in CoL Iraq
Halticus henschii Reuter, 1888 View in CoL Italy
Halticus insularis Usinger, 1946 Mariana Is. View in CoL ; Guam; Korea
Halticus intermedius Uhler, 1904 View in CoL USA
Halticus luteicollis ( Panzer, 1804) View in CoL * Europe; Mediterranean
Halticus macrocephalus Fieber, 1858 View in CoL Mediterranean
Halticus major Wagner, 1951 View in CoL Europe
Halticus obscurior Kerzhner & Muminov, 1974 Asiatic View in CoL Russia
Halticus puncticollis Fieber, 1870 View in CoL Montenegro
Halticus pusillus ( Herrich-Schäeffer, 1835) View in CoL * Palaearctic
Halticus rugosus Reuter, 1894 View in CoL Syria
Halticus saltator ( Geoffroy, 1785) View in CoL Europe
Halticus tibialis Reuter, 1891 View in CoL * Africa; South-East Asia, South Pacific
Biology and host plant associations: Limited host records exist for Halticus . Halticus apterus and H. beganus have both been collected from the family Fabaceae ( Vicia striata and Phaseolus vulgaris , respectively). Halticus beganus has also been collected from rotten oak ( Fagaceae ) ( Henry & Smith, 1979). Halticus pusillus is said to live on and beneath Galium verum (Rubiaceae) ( Wagner, 1973) ( Table 1). Remarks: Most recent definitions of Halticus are broad, inclusive of great morphological heterogeneity (e.g. Wagner, 1973), and pay little regard to genitalic features. This is the first study to account for the morphological diversity exhibited across species of Halticus , and to examine this in a phylogenetic context. As a consequence we have subdivided Halticus sensu lato into three genera. Based primarily on male and female genitalic characters, and supported by our phylogeny, we remove ten species examined that do not follow the model of the type species, H. apterus (left paramere L-shaped with apically curved apophysis; ductus seminis short; secondary gonopore elongate and scoop-shaped, endosoma obsolete and without spicules; sclerotized rings large and subcontiguous). H. bractatus , H. canus , H. chrysolepis , H. minutus , and H. spegazzini are removed, and along with both species of Cafayatina , form the resurrected genus Microtechnites . Similarly, the Asian species H. bicoloratus , H. comitans , H. maculipes , H. niger , and H. fuscous are all transferred to Ectmetopterus .
We have not seen all species of Halticus , and it is therefore possible that with further examination other species may yet be removed from the genus.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Halticus
| Tatarnic, Nikolai J. & Cassis, Gerasimos 2012 |
Halticiellus
| Carvalho JCM 1958: 12 |
| Villiers A 1952: 186 |
Halticocoris
| Carvalho JCM 1958: 13 |
| Douglas JW & Scott J 1865: 478 |
Halticus
| Cassis G & Gross GF 1995: 186 |
| Linnavuori RE 1994: 5 |
| Henry T 1983: 610 |
| Kelton LA 1980: 194 |
| Alayo DP 1974: 15 |
| Schuh RT 1974: 29 |
| Wagner E 1973: 6 |
| Maldonado CJ 1969: 57 |
| Kerzhner IM 1964: 968 |
| Wagner E 1961: 48 |
| Southwood TRE & Leston D 1959: 244 |
| Kelton LA 1959: 33 |
| Carvalho JCM 1958: 12 |
| Carvalho JCM 1956: 51 |
| Carvalho JCM 1955: 65 |
| Wagner E 1952: 95 |
| Carvalho JCM & Leston D 1952: 245 |
| Kiritshenko AN 1951: 127 |
| Slater JAS 1950: 45 |
| Zimmerman EC 1948: 199 |
| China WE 1943: 268 |
| Hsiao TY 1942: 253 |
| Knight HH 1941: 17 |
| Stichel W 1933: 235 |
| Blatchley WS 1926: 800 |
| Knight HH 1923: 498 |
| Van Duzee EP 1916: 211 |
| Poppius B 1914: 84 |
| Oshanin B 1910: 802 |
| Reuter OM 1910: 147 |
| Kirkaldy GW 1906: 130 |
| Hueber T 1906: 2 |
| Distant WL 1904: 479 |
| Saunders E 1892: 267 |
| Reuter OM 1891: 17 |
| Atkinson ET 1890: 117 |
| Reuter OM 1875: 23 |
| Reuter OM 1875: 86 |
| Saunders E 1875: 287 |
| Fieber FX 1861: 69 |
| Fieber FX 1858: 312 |
| Spinola MM 1850: 43 |
| Burmeister HCC 1835: 277 |
| Hahn CW 1833: 113 |
Eurycephala
| Carvalho JCM 1958: 12 |
| Kirkaldy GW 1906: 170 |
| Reuter OM 1891: 17 |
| Spinola MM 1840: 191 |
| Blanchard E 1840: 140 |
| Brulle MA 1836: 140 |
| Laporte FL 1832: 36 |
Astemma
| Amyot CJB & Serville A 1843: 284 |
| Westwood JO 1840: 121 |
| Latreille PA 1829: 199 |