
Pseudonereis, KINBERG, 1865
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2007.00289.x |
|
persistent identifier |
https://treatment.plazi.org/id/03E7C033-FFB1-FFAA-F55C-E7A127CEF95D |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudonereis |
| status |
|
PSEUDONEREIS KINBERG, 1865 View in CoL
Pseudonereis Kinberg, 1865: 174 View in CoL . – Fauchald, 1977: 90.
Type species. Pseudonereis gallapagensis Kinberg, 1865 designated by Hartman (1949).
Diagnosis: Prostomium with entire anterior margin, one pair of antennae, one pair of biarticulated palps with conical palpostyles, four pairs of tentacular cirri with distinct cirrophores. Two pairs of eyes. One apodous anterior segment, greater than length of chaetiger 1. Maxillary ring of pharynx with paragnaths, arranged in discrete areas, Areas II–IV p-bars and conical paragnaths arranged in regular comb-like rows. Oral ring paragnaths present, Area V conical paragnaths present or absent; Area VI conical paragnaths present or absent, shield-shaped bars present or absent; Area VII–VIII p-bars and conical paragnaths present. Dorsal notopodial ligule markedly elongate and markedly broader on posterior chaetigers. Prechaetal notopodial lobe present or absent. Dorsal cirrus terminally attached to dorsal notopodial ligule on posterior chaetigers (subterminally in P. anomala ). Neuropodial superior lobe present (absent in P. palpata and P. pseudonoodti ). Neuropodial postchaetal lobe absent or present, at least on some anterior chaetigers. Notoaciculae absent from chaetigers 1 and 2. Notochaetae homogomph spinigers present, homogomph falcigers present or absent. Neurochaetae, dorsal fascicle: heterogomph spinigers present or absent, homogomph spinigers present or absent, heterogomph falcigers present, blades serrated. Neurochaetae, ventral fascicle: heterogomph spinigers and heterogomph falcigers present.
Remarks: Bakken & Wilson (2005) recently redefined the genus based on phylogenetic analyses of the subfamily Nereidinae . In the description above minor changes are included (highlighted in bold). Position of the dorsal cirrus in P. anomala was coded as terminally attached in their analyses. Re-examination of types and additional material examined in this study shows that the position of the dorsal cirrus is subterminal. In P. multisetosa the dorsal notopodial ligule is clearly not elongated in posterior chaetigers, a character that should be verified when more material is found, as the single known specimen is probably a juvenile.
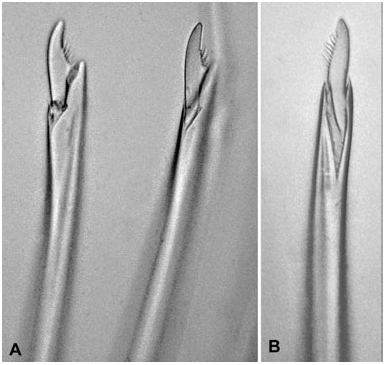
Presence of conical paragnaths arranged in regular comb-like rows in Areas II–IV ( Fig. 1A–C View Figure 1 ) is unique to Pseudonereis species. Presence of a shield-shaped paragnath in Area VI, here described for the first time, is also shared in Pseudonereis species except P. palpata (possesses both a bar and conical paragnaths in Area VI), and P. anomala and P. multisetosa (cones only). A comparison for all taxa is presented in Table 1.
PSEUDONEREIS ANOMALA GRAVIER, 1901 View in CoL
( FIGS 2 View Figure 2 AND 3 View Figure 3 )
Pseudonereis anomala Gravier, 1901: 191–197 View in CoL , TII, pl. XII figs 50,51,52, text figs 194–202. – Hutchings & Turvey 1982: 141–142. – Hutchings & Glasby 1985: 108–109. – Hylleberg et al. 1986: 13–14, fig. 7.
Nereis (Pseudonereis) rottnestiana Augener, 1913: 184–187 View in CoL , Taf. III, fig. 46, text-fig. 20a–c.
Nereis nichollsi Kott, 1951: 93–95 View in CoL , fig. 2a–k.
Pseudonereis rottnestiana View in CoL forma seriodentata Hartmann-Schröder, 1979: 118–119, figs 216–219. – 1980: 61.
Material examined: Lectotype MNHN TYPE 1466 (1), paralectotypes, Djibouti 1897 (n29 1897), coll. Coutière, H., MNHN POLY TYPE- 423 (12); syntypes, Djibouti 1897, coll. Coutière, H., POLY TYPE- 421 (2); Khark, off East side, Iran, 6/31937, coll. G. Thorson, ZMUC-POL-1802 (13); Cape de Couedic, Kangaroo Island, SA, algal holdfast, exposed reef, 4.iii.1979, coll. Hutchings & Butler, AM W18310 (98); Cape de Couedic, Kangaroo Island, SA, algal holdfast, exposed reef, 4.iii.1979, coll. P. Hutchings W18311 (47); Cape de Couedic, Kangaroo Island, SA, algal holdfast, exposed reef, 4.iii.1979, coll. P. Hutchings AM W18312 (6); Mastan Pt. America River, Kangaroo Island, clumps of sponge in fast flowing channel, 2.iii.1979, coll. P. Hutchings, AM W18313 (1); Pelsart Group, H. Abrolhos Islands, WA, rock wastings of reef, 1.ix.1947, coll. P. Kott, AM W18574 (1); Plantation Point, NSW 27. VI.1981, AM W25645 (12); North West Solitary Island, coralline algae, 25.vi.1992, coll. P.B. Berents, AM W25646 (1); Broome, probe 6, 9.ix.1975, coll. G. Hartmann-Schröder (5) ZMH P-16564; Port Hedland, probe 34, 30.ix.1975, coll. G. Hartmann-Schröder (1), ZMH P-16565. Nereis (Pseudonereis) rottnestiana , syntype, Rottnest, ZMH V-7929 (1). Nereis nichollsi , holotype Rottnest Island, Point Peron, Abrolhos, WA, from weed rock outer edge of reef, coll & id Pat Kott 1946−49, AM W7036 (1); paratypes Rottnest Island, Point Peron, Abrolhos, WA, from weed rock outer edge of reef, coll & id Pat Kott 1946−49, AM W7037 (31). Size range of material examined 48–86 chaetigers (23–70 incomplete specimens), 10–46 mm long (11–34 incomplete specimens), 1–3.5 mm body width.
Description: Frontal antennae present, 1 pair, cirriform. Prostomium with entire anterior margin. Eyes present, 2 pairs. One apodous anterior segment, greater than length of chaetiger 1. Tentacular cirri with distinct cirrophores, longest tentacular cirri extend back to chaetiger 4–9. Jaws with dentate cutting edge, brown-black, plate-like serrated with 4–5 teeth. Maxillary ring of pharynx with paragnaths arranged in discrete areas, Areas II–IV arranged in regular comb-like rows. Area I = 1–3 conical paragnaths (4 in two specimens, n = 39); Area II = 11–31 p-bar paragnaths in 3– 4 rows; Area III = 30–72 p-bar paragnaths in 4 rows; Area IV = 20–52 p-bar paragnaths in 4 rows and additional cones towards the jaws. Area V and VI present as distinct groups. Area V paragnaths absent; VI = 3– 15 conical paragnaths, bars absent; VII–VIII = 10–24 conical paragnaths, arranged in one row with similar sized cones, or with two rows where the second row has slightly smaller cones interspersed in the first row or posterior to the first.
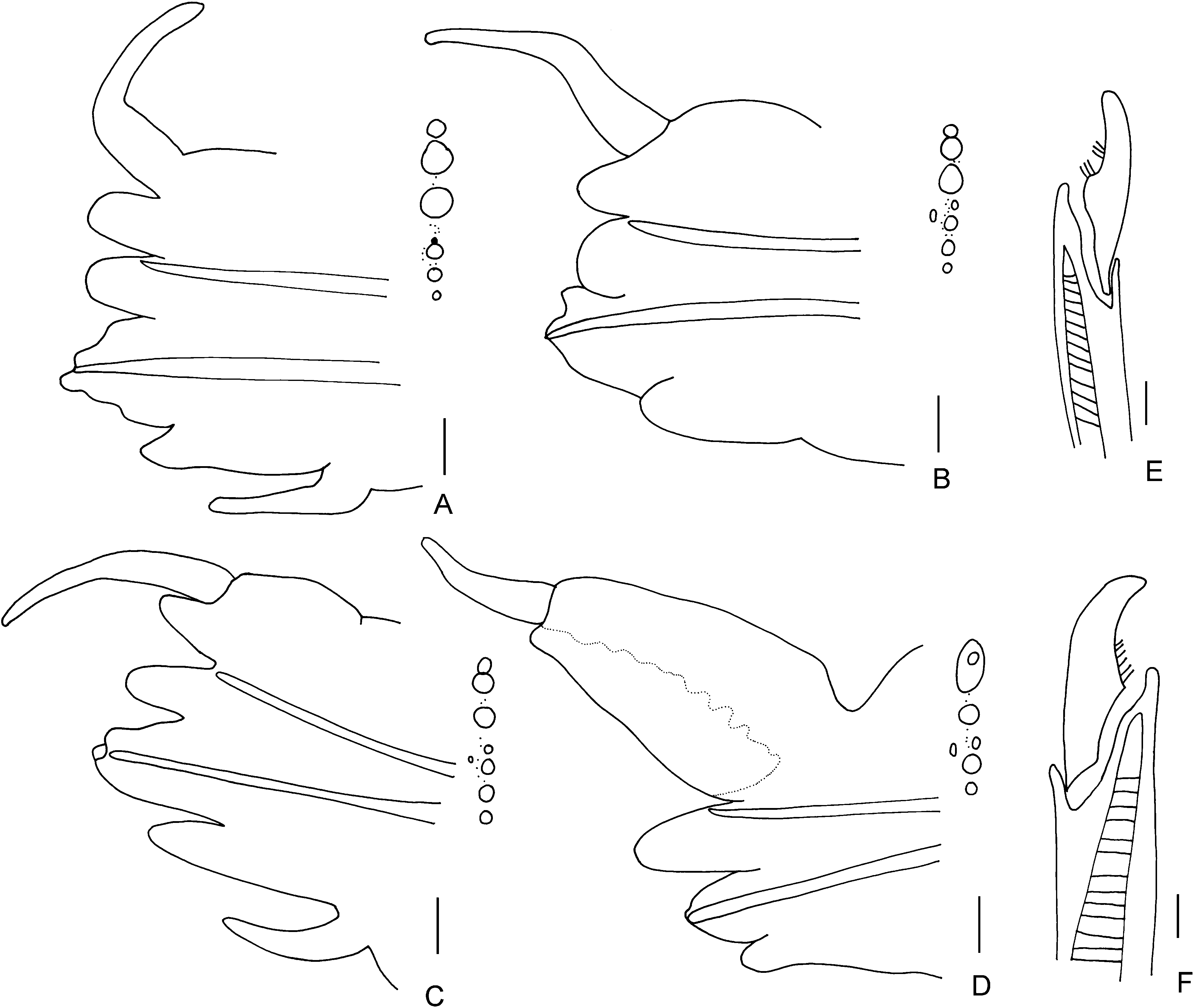
Notopodium with dorsal notopodial ligule rounded, as long as ventral notopodial ligule on anterior ( Fig. 3A View Figure 3 ) and mid-body chaetigers ( Fig. 3B View Figure 3 ), markedly elongate and broader on posterior chaetigers ( Fig. 3C, D View Figure 3 ). Prechaetal notopodial lobe absent; acicular process absent. Dorsal cirrus basally attached to dorsal notopodial ligule in anterior chaetigers ( Fig. 3A, B View Figure 3 ), subterminally attached on posterior chaetigers ( Fig. 3D View Figure 3 ). Dorsal cirrus simple, lacking basal cirrophore, 3–4 times as long as ventral notopodial ligule at chaetigers 10–20 ( Fig. 3B View Figure 3 ). Ventral notopodial ligule rounded, slender with a rounded tip in posterior chaetigers.
Neuropodial inferior lobe prominent in anterior chaetigers; a small superior lobe present throughout; neuropodial postchaetal lobe present throughout as a low lobe, not projecting beyond end of acicular ligule, most prominent and drawn out to a triangular tip in posterior chaetigers. Ventral neuropodial ligule similar in length to acicular neuropodial ligule throughout, rounded in anterior chaetigers, slender in posterior. Ventral cirrus similar in length or slightly shorter than acicular ligule.
Notochaetae: homogomph spinigers ( Fig. 3F View Figure 3 ) and falcigers present, falcigers first appear from about chaetiger 30, homogomph falcigers serrated ( Figs 2B View Figure 2 , 3H View Figure 3 ). Neurochaetae, dorsal fascicle: homogomph spinigers and heterogomph falcigers present, blades serrated. Neurochaetae, ventral fascicle: heterogomph spinigers present, one or two heterogomph spinigers present from about chaetiger 40 to 50, homogomph spinigers absent, heterogomph falcigers with short blades in anterior and posterior chaetigers present ( Figs 2A View Figure 2 , 3G View Figure 3 ).
Anal cirri reaching back 8–10 chaetigers. Epitokal modification observed in two specimens ( MNHN Poly Type-421), both females filled with large eggs, parapodial modification ( Fig. 3E View Figure 3 ) starting from chaetiger 18.
Remarks: A lectotype is designated from the syntype series ( MNHN POLY TYPE- 423). The lectotype is a complete specimen measuring 41 mm for 84 chaetigers, 2.5 mm wide. The other 11 specimens from the syntype series are designated paralectotypes. The original description was presented by Gravier (1901) with text figures, but the accompanying figures on plates were printed in an earlier work also on polychaetes from the Red Sea ( Gravier, 1900).
The digitiform tip of the dorsal notopodial ligule in posterior chaetigers is obvious and the subterminal position if the dorsal cirrus is very clear. In most specimens the tip of the dorsal ligule is a digitiform protrusion, but this is subject to some variation as it might be a curved tip. This is the case for the syntype of P. rottnestiana , but there is gradual variation over several specimens from the same population (in the paratype series of Nereis nichollsi ). The protruding tip of the dorsal ligule and the dorsal cirrus in a subterminal position is different from the other taxa in the genus, except P. multisetosa , which have the dorsal cirrus in a terminal position. This new information corrects an error in Bakken & Wilson (2005), who coded the dorsal cirrus as terminally attached in posterior chaetigers for this taxon. In the neuropodia a small postchaetal lobe is present. Although it is prominent, typically in posterior chaetigers with a triangular tip, it might be difficult to see especially in smaller specimens (body width <1 mm).
Pseudonereis anomala differs from other Pseudonereis taxa by the presence of notopodial homogomph falcigers and presence of conical paragnaths in Area VI, also present in P. multisetosa . Pseudonereis anomala can be distinguished from P. multisetosa by presence of homogomph falcigers in dorsal fascicle of neuropodia in that the latter.
Pseudonereis rottnestiana View in CoL was described from Rottnest Island, Western Australia ( Augener, 1913), the same locality from where Kott (1951) described Nereis nichollsi View in CoL . Augener (1913) stated he had two specimens, one small and one larger. The original material was not labelled as types, but according to the measurements given in his description the larger one is identical to a specimen examined here (ZMH V-7929). The specimen is in very good condition but most chaetae are broken; no single notopodial chaeta was observed. Heterogomph spinigers in the neuropodial fascicle were observed in a few posterior chaetigers. Augener (1913) did not include in his description chaetae other than from a ‘middle parapodium’, did not mention notopodial homogomph falcigers, and he did not mention chaetae in more posterior chaetigers. Absence of notopodial homogomph falcigers in P. rottnestiana View in CoL has been given as the only difference between the two similar P. anomala View in CoL and P. rottnestiana View in CoL ( Fauvel, 1932; Hartman, 1954). Augener’s only comparison of P. rottnestiana View in CoL with P. anomala View in CoL was his statement that the two species differed in posterior parapodia, which they indeed do judged from this single specimen. However, there are variations in the shape of the dorsal notopodial ligule in this species, even within a population.
Hutchings & Glasby (1985) discussed the number of conical paragnaths in Area VI and their arrangement in one or two rows in an arc. Variation in paragnath numbers was also discussed by Hutchings & Turvey (1982) (type specimens of N. nichollsi View in CoL ). Although few specimens were examined here, it seems that paragnath numbers are consistent within populations but show minor variations between populations (Table 2). Other morphological differences between the populations are not observed. This is also consistent with two rows of conical paragnaths in Area VI, along with other characters described and illustrated in specimens from Thailand by Hylleberg et al. (1986).
Hartmann-Schröder (1979, 1980) reported two different forms, published as ‘ Pseudonereis rottnestiana forma seriodentata ’ and ‘ Pseudonereis rottnestiana forma costatodentata ’. Describing forms or other infrasubspecific names is nomenclatorially invalid and does not constitute description of a new taxon (ICZN, 1999). It is likely that this represents an accidental publication of manuscript notes. It is evident from the specimens in question (ZMH P-16564; P-16565) that the form Hartmann-Schröder (1979) described as ‘ seriodentata ’ represents specimens of P. anomala , while her form ‘ costatodentata ’ represents specimens of P. trimaculata (see remarks for this taxon).
Distribution: Type locality Djibouti, Gulf of Aden, Arabian Sea; known from the Arabian Sea, Persian Gulf, around Australia except in the northern part, Thailand ( Hylleberg et al., 1986), Hainan Island and Xisha Island, China ( Wu, Sun & Yang, 1985). The depth distribution is intertidal and in shallow water.
PSEUDONEREIS ATOPODON CHAMBERLIN, 1919 View in CoL
( FIG. 4 View Figure 4 )
Pseudonereis atopodon Chamberlin 1919: 228–229 View in CoL , pl. 35, figs 3–5.
Material examined: Holotype, Nomuka , Tonga Island 12.ii.1899, intertidal, USNM 19467 About USNM ; paratypes, Nomuka, Tonga Island 12.ii.1899, intertidal, USNM 19468 About USNM (2). Size range of material examined 87–92 chaetigers, 35–44 mm long, 1 mm wide.
.
examined’ VIII – VII 13– 15 ± 1.0 13.5 (
) – 19 16 ± (17.3) 1.3 10 16 – ± () 2.0 13.3 19 – 14 ± 16.0 2.2) (– 17 10 ± 14.2 2.3) (– 11 14 ± 1.3 13.4 () Material
±
) 2.5) 1.1 1.9) 1.7) 1.5) 0.5)
in
‘ series R VI 9 15 – 11.5 (9 – 6 ± (6.9 5 10 – ± 7.4 (11 7– ± 9.5 (– 9 4 ± 5.9 (4 5 – ± (4.4 given type
are the ± 1.7
) 1.4) 2.3)) 2.2) 0.9) 0.5 data from VI L 13 – 11.5 9 – ± 6.9 – 12 ± 8.1 12 – ± 9.3 – 7 ± 5.9 6 – ± 4.4 location syntypes V 0 9 (5 0 (6 0 (0 7 (5 0 (3 0 (
Exact. localities represent R IV – 33 27 ± 2.9 28.8) (28 51 – ± 36.9 6.9 () 38 27– ± 32.6) 4.0 (48 – 38 ± 44.5 4.4 () 24 – 42 ± () 33.0 6.4 38 26 – ± 4.8 33.6) (
different six
Djibouti from IV L 20 – 33 ± 26.0 6.1) – 29 52 ± 6.3) 37.9 – 27 38 ± 4.5 31.3) 35 – 49 ± 6.1) 41.8 27 – 46 ± 34.3 6.1) 40 32 – ±) 36.2 3.9
of specimens Specimens 3.2)
((
) 8.2
(
) 5.6) 10.0
(
6.8
(
) 4.3
(
). pharynx III 36 30 – ± (32.3 37 63 – ± (48.2 – 53 37 ± 43.1 (– 51 72 ± (60.3 – 56 36 ± 43.6 (36 46 – ± 42.0 (examined the 2.4)) 3.6 3.4)) 2.5 2.1) 4.3) of ± ± ± ± ± ± from area II R 11 – 16 13.5 (17 – 29 21.8 (14 – 23 18.7 (20 – 26 23.3 (– 20 14 18.0 (24 – 14 19.2 ( anomala each for 1.8) ±) 4.7 ±) 1.6) 1.4 2.2) ±) 3.7
Pseudonereis ± SD mean
) ( II L 12 16 – ± (14.0 – 28 15 0.7 20.8 () – 18 22 0.4 20.1) (23 – 26 ± 1.3 () 25.0 20 – 14 ± 17.2 0.5) – 14 23) (20.0 1.1 for and 3 ± 3 ± 4 ± 2 ± 4 ± statistics as range n I 4 1 9 1 – (1.2 – 2 7 (2.1 – 1 4 2.5 (1 9 – 1.7 (1 – 5 (2.2 Paragnath given are
, Point Australia, Island Australia
,
Western Western
Table
2
.
Numbers Locality Plantation, NSW Kangaroo South Dampier Australia, Broome Australia Djibouti, Iran Khark Description: Frontal antennae present, 1 pair, cirriform. Prostomium with entire anterior margin, longer than wide. Eyes present, 2 pairs. One apodous anterior segment, greater than length of chaetiger 1. Tentacular cirri with distinct cirrophores, longest tentacular cirri extend back to chaetiger 4. Jaws with dentate cutting edge, brown, plate-like with 4 teeth. Maxillary ring of pharynx with paragnaths arranged in discrete areas, Areas II–IV arranged in regular comb-like rows. Area I = 1; Area II = 37–49 p-bar paragnaths in 5 rows; Area III = 56 p-bar paragnaths in 5 rows; Area IV = in 4–5 rows, cones towards jaws (impossible to see details due to small size or broken), bar-shaped paragnaths or p-bars not observed. Area V and VI present as distinct groups. Area V = 0–1 conical paragnaths, when present a large cone; Area VI = 1 large shield-shaped bar with a pointed tip (triangular) present; Area VII–VIII = 14–24 paragnaths with an appearance like p-bars, similar in size.
Notopodium with dorsal notopodial ligule short, rounded as long as ventral notopodial ligule in anterior chaetigers ( Fig. 4A View Figure 4 ), markedly elongate and markedly broader on posterior chaetigers ( Fig. 4D View Figure 4 ). Prechaetal notopodial lobe absent, acicular process absent. Dorsal cirrus simple, lacking basal cirrophore, 3 times as long as ventral notopodial ligule at chaetigers 10–20 ( Fig. 4B View Figure 4 ), basally attached to dorsal notopodial ligule in anterior and mid-body chaetigers, terminally attached to dorsal notopodial ligule on posterior chaetigers ( Fig. 4D View Figure 4 ), commencing at about chaetiger 60–65.
Neuropodial inferior lobe prominent in anterior chaetigers, a small superior lobe present; neuropodial postchaetal lobe present throughout ( Fig. 4C View Figure 4 ), a low, rounded, flattened lobe level with acicular ligule or lower. Ventral neuropodial ligule rounded up to half length of acicular neuropodial ligule, similar throughout. Ventral cirri short.
Notochaetae: homogomph spinigers. Neurochaetae, dorsal fascicle: homogomph spinigers present; heterogomph falcigers present throughout, blades serrated. Neurochaetae, ventral fascicle: heterogomph spinigers present from about chaetiger 30, heterogomph falcigers with short blade present throughout ( Fig. 4E, F View Figure 4 ).
Anal cirri reaching back 4–5 chaetigers.
Remarks: The neuropodial postchaetal lobe is most prominent in mid-body chaetigers ( Fig. 4C View Figure 4 ). Pseudonereis atopodon is very similar to P. gallapagensis , the only differences being higher number of paragnaths in Area II in P. atopodon (37–49) than in P. gallapagensis (17–20), and the length of the ventral neuropodial ligule being up to 0.5 times as long as neuropodial acicular ligule in P. atopodon , compared with as long as acicular ligule in P. gallapagensis . Pseudonereis atopodon is also very similar to P. variegata , being distinguished by the same relationship of the neuropodial ventral ligule as to P. gallapagensis , and in the length of the dorsal cirrus being longer in P. atopodon than in P. variegata ( Table 1).
Obviously P. atopodon is very similar to P. gallapagensis and P. variegata and the identity of the three should be taken into account when more material is available. Differences in paragnath numbers are minor and have not been analysed statistically (more material is required). Of the three specimens examined it was possible in only one to count paragnath numbers for Area III, and none for Area IV.
Chamberlin (1919) stated this taxon was similar to P. gallapagensis but did not give a further justification.
Distribution: Nomuka, Tonga Island (type locality), intertidal ( Chamberlin, 1919).
PSEUDONEREIS CORTEZI ( KUDENOV, 1979) View in CoL
( FIGS 5 View Figure 5 , 6 View Figure 6 )
Neanthes cortezi Kudenov, 1979: 118–120 View in CoL , fig. 2a–h; de León-González & Solís-Weiss 2000: 554–555.
Material examined: Paratypes, Punta La Cholla , Sonora, Mexico, from Tetraolita squamosa tests, x.1976, NHMLAC Poly 1344 (23). Size range of material examined 65–90 chaetigers (37–69 incomplete specimens), 10–37 mm long (13–23 incomplete specimens), 0.5–1 mm wide .
Description: Frontal antennae present, 1 pair, cirriform. Prostomium with entire anterior margin, longer than wide. Eyes present, 2 pairs. One apodous anterior segment, greater than length of chaetiger 1. Tentacular cirri with distinct cirrophores, longest tentacular cirri extend back to chaetiger 3–5. Jaws with dentate cutting edge, dark brown with 9–11 teeth. Maxillary ring of pharynx with paragnaths, arranged in discrete areas, Areas II–IV arranged in regular comb-like rows. Area I = 4–6 conical paragnaths in longitudinal rows; Area II = 17–31 p-bars and conical paragnaths in 4–5 rows in a triangular patch; Area III = 33–54 p-bars and conical paragnaths in 5 rows; Area IV = 31–55 p- bars and conical paragnaths in 4– 5 long and 3–4 short rows, shorter rows being closer towards the jaws, and 1–3 p-bars next to the jaws. Area V and VI present as distinct groups. Area V = 8– 15 conical paragnaths arranged in a triangular pattern ( Fig. 5A View Figure 5 ); Area VI = 1 large shield-shaped bar present, in some specimens with a distinct pointed tip, cones absent; Area VII–VIII = 62–87 conical paragnaths similar in size forming a broad band, with interspersed p-bars ( Fig. 5B View Figure 5 ).
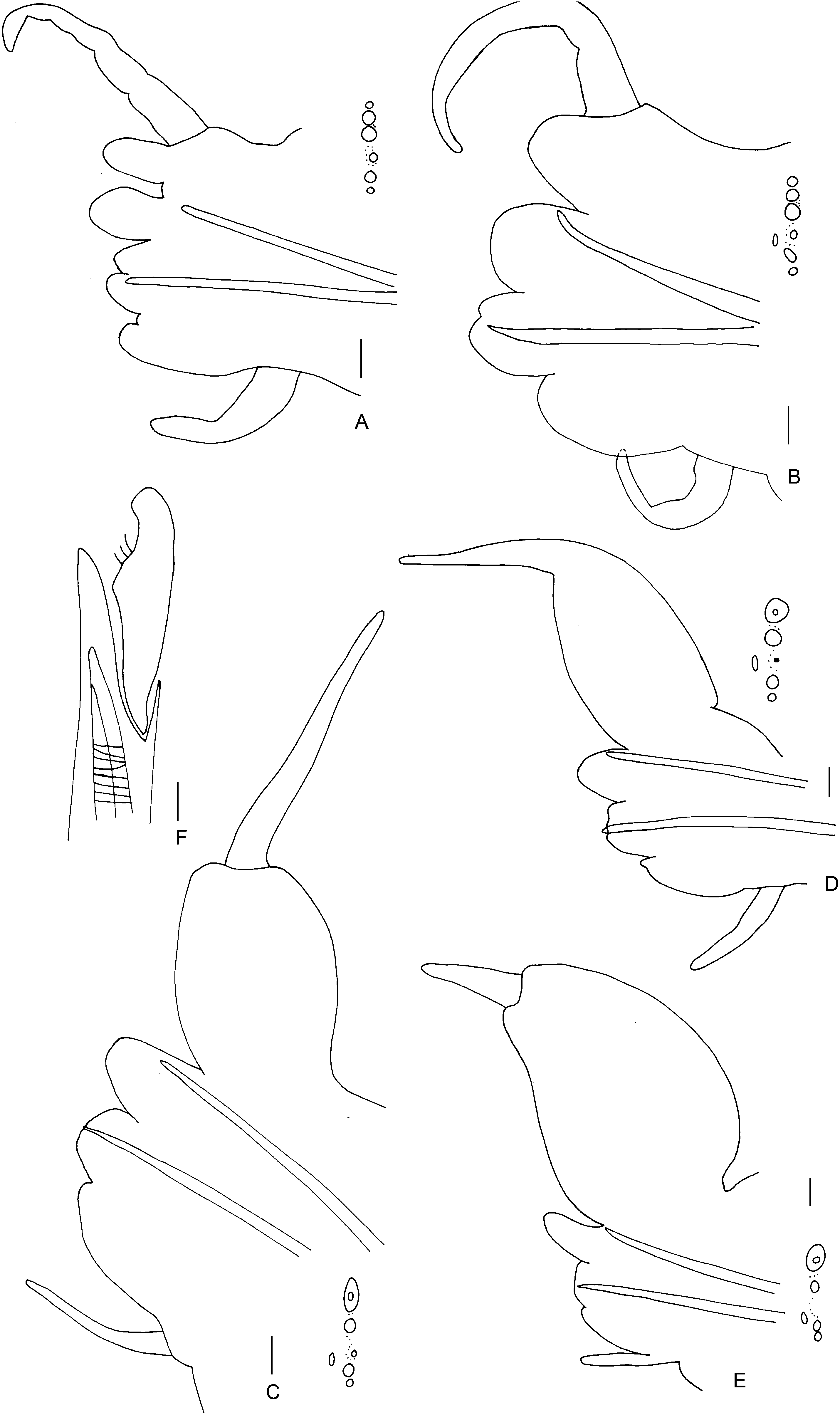
Notopodium with dorsal notopodial ligule short and rounded as long as ventral notopodial ligule in anterior chaetigers ( Fig. 6A, B View Figure 6 ), markedly elongate and markedly broader on posterior chaetigers, starting from about chaetiger 50–55 ( Fig. 6E, F View Figure 6 ). Prechaetal notopodial lobe absent, acicular process absent. Dorsal cirrus simple, lacking basal cirrophore, 2.5 times ventral notopodial ligule at chaetiger 10–20 ( Fig. 6B View Figure 6 ), basally attached to dorsal notopodial ligule in anterior chaetigers, subterminally attached from about chaetiger 35–40 ( Fig. 6D View Figure 6 ), terminally attached from about chaetiger 50–55.
Neuropodial inferior lobe poorly developed; a small superior lobe gives the acicular ligule a bilobed appearance ( Fig. 6C View Figure 6 ), restricted to the first about 40 chaetigers. Neuropodial postchaetal lobe absent. Ventral neuropodial ligule poorly developed, less than 0.5 times as long as acicular ligule, similar to posterior chaetigers, reduced to a papilla ( Fig. 6F View Figure 6 ) in the last about 40 chaetigers. Ventral cirri 0.5 times shorter than ventral ligule.
Notochaetae: homogomph spinigers. Neurochaetae, dorsal fascicle: homogomph spinigers present; heterogomph falcigers present ( Fig. 6G View Figure 6 ) throughout, blades serrated. Neurochaetae, ventral fascicle: a single heterogomph spiniger present from chaetiger 5–10, heterogomph falcigers with short blade present throughout.
Pygidium with multiple incisions, anal cirri reaching back 3–4 chaetigers. Pigmentation in most specimens part of the palps and the prostomium dark brown, except for a small unpigmented lance-shaped patch in the centre ( Fig. 5A View Figure 5 ; Kudenov, 1979: fig. 2a).
Remarks: The original description ( Kudenov, 1979) stated that a single conical paragnath was present in Area VI. This is, however, a large, laterally flattened, high and triangular shield-shaped bar as in most other Pseudonereis species. In some specimens this bar has a distinct point skewed towards one side. In Area III the conical paragnaths are more spaced ( Fig. 5B View Figure 5 ) as is usual in other species with comb-shaped rows, although still tight rows. In Areas II and IV the closely comb-shaped rows appear as commonly observed in Pseudonereis species. It is very difficult to see in the specimens examined here where the notopodial dorsal ligule ends and where the dorsal cirrus starts, and it is likely the expansion in breadth and length of the dorsal ligule is quite small so that the cirrus is rather long and has a wide base. In most other taxa the dorsal cirrus is attached to the tip of the dorsal ligule and is usually 1/4–1/6 of the length of the ligule. In this taxon it appears that the dorsal cirrus comes to a major part of the dorsal ligule. This question must be left unanswered until more material, preferably live, is available.
Pseudonereis cortezi is distinguished from the other taxa by the absence of a neuropodial postchaetal lobe, which is usually prominent in posterior chaetigers, and by the high number of paragnaths in Area V (8– 15) ( Fig. 5A View Figure 5 ) while the other taxa have 0–3. Pseudonereis cortezi most closely resembles P. pseudonoodti but can be distinguished from the latter by presence of a neuropodial superior lobe in anterior and mid-body chaetigers.
Distribution: Type locality Bahia Cholla, Puerto Peñascola, northern Gulf of California
PSEUDONEREIS GALLAPAGENSIS KINBERG, 1865 View in CoL
( FIG. 7 View Figure 7 )
Pseudonereis gallapagensis Kinberg, 1865: 174 View in CoL . – Hartman 1949: 68–69. – Hartmann-Schröder 1962b: 432–434. – Fauchald 1977: 32–33.
Pseudonereis formosa Kinberg, 1865: 174 View in CoL .
Material examined: Syntype, Indifatigable Island , Galapagos, Eugenie Exp. 1851−53, stn 873, SMNH Type-452 (1); Valparaiso, Chile, Eugenie Exp. 1851− 53, stn 500, SMNH 37896 View Materials (1) ; Chincha Island , Peru, Eugenie Exp. 1851−53, stn 531, SMNH 37897 View Materials (1) ; Chincha Island , Peru, Eugenie Exp. 1851−53, stn 531, SMNH 37898 View Materials (1); Syntype Pseudonereis formosa, Honolulu , Hawaii, Eugenie Exp. 1851−53, stn 1081− 91, depth 45 m, SMNH Type-5908 (1). Size range of material examined 43–73 chaetigers (incomplete specimens), 26–47 mm long, 2–2.5 mm wide .
Description: Frontal antennae present, 1 pair, cirriform. Prostomium with entire anterior margin, wider than long. Eyes present, 2 pairs. One apodous anterior segment, greater than length of chaetiger 1. Tentacular cirri with distinct cirrophores, longest tentacular cirri extend back to chaetiger 3–4. Jaws with dentate cutting edge, dark brown, with 4–5 teeth. Maxillary ring of pharynx with paragnaths, arranged in discrete areas; Areas II–IV arranged in regular comb-like rows. Area I = 1 conical paragnath; Area II = 17–20 pbar paragnaths in four rows; Area III = 51–68 p-bar paragnaths in 4 rows; Area IV = 38–57 including pbar paragnaths in 4 rows, around 15 cones towards jaws and 2–4 p-bars next to the jaws. Oral ring paragnaths present, Area V and VI present as distinct groups. Area V = 1 conical paragnath, Area VI = 1 large triangular shield-shaped bar present, cones absent; Area VII–VIII = 17–20 in two rows, anterior row with cones, posterior with p-bars, forming a single band of paragnaths.
Notopodium with dorsal notopodial ligule short and rounded ( Fig. 7A, B View Figure 7 ) markedly elongate and markedly broader on posterior chaetigers ( Fig. 7C–E View Figure 7 ), expansion from about chaetiger 15. Prechaetal notopodial lobe absent, acicular process absent. Dorsal cirrus simple, lacking basal cirrophore, 3 times ventral notopodial ligule at chaetiger 10–20 ( Fig. 7B View Figure 7 ), basally attached in anterior chaetigers, terminally attached to dorsal notopodial ligule on posterior chaetigers, commencing at about chaetiger 15. Ventral notopodial ligule rounded as long as neuropodial acicular ligule in anterior chaetigers, longer than, protruding, posteriorly.
Neuropodial inferior lobe prominent in anteriormost chaetigers only, a small superior lobe present ( Fig. 7A, B View Figure 7 ). Neuropodial postchaetal lobe present throughout, projecting level with acicular ligule as a low rounded flattened lobe, with a pointed tip in posterior chaetigers. Ventral neuropodial ligule of anterior chaetigers similar in length to acicular neuropodial ligule, up to half length of acicular neuropodial ligule in posterior chaetigers. Ventral cirri as long as acicular ligule throughout.
Notochaetae: homogomph spinigers. Neurochaetae, dorsal fascicle: homogomph spinigers present; heterogomph falcigers present on anterior ( Fig. 7F View Figure 7 ) and posterior chaetigers, blades serrated. Neurochaetae, ventral fascicle: heterogomph spinigers, usually a single, present in posterior chaetigers, heterogomph falcigers with short blade present throughout.
Pygidium with multiple, ventral incisions, anal cirri reaching back 4–5 chaetigers (observed in one specimen).
Remarks: The description above is based on the syntype of P. gallapagensis ( SMNH Type-452) and the syntype of P. formosa ( SMNH Type-5908), the latter being the more complete specimen, which is also used for illustrations ( Fig. 7 View Figure 7 ). The type of P. gallapagensis consists of one anterior end and three other fragments from the same specimen in poor condition. It is very difficult to get details from the different pieces, as they are flattened and the parapodia are a little deformed. Patterns of paragnaths in the different areas of the pharynx are possible to see but it is not possible to count numbers. One heterogomph spiniger was observed in ventral fascicle in mid-body and posterior chaetigers; heterogomph spinigers were not observed in the type specimen of P. formosa . Otherwise these specimens are very similar and are judged to be conspecific. Hartman (1949) stated P. formosa was a synonym of P. gallapagensis after examining type material of both specimens and the material was labelled accordingly. Re-examination of the material for this study revealed an original label in the vial with the specimens from Honolulu reading P. formosa in faded yellow ink. This specimen is clearly the type specimen of P. formosa ( SMNH Type-5908).
No differences were observed in the material examined from Galapagos, Hawaii, Peru and Chile listed above, but specimens are not in good condition. Hartman (1949) concluded that the specimens identified as Neanthes variegata by Kinberg (1865) ( SMNH 37897, SMNH 37898) can be referred to P. gallapagensis and I agree.
Pseudonereis gallapagensis View in CoL is most similar to P. variegata View in CoL and P. atopodon View in CoL , and can be distinguished from P. variegata View in CoL by the longer dorsal cirrus, ventral neuropodial ligule up to half as long as acicular ligule in posterior chaetigers compared with as long as in P. variegata View in CoL , and by having fewer paragnaths in Area IV. In the literature (e.g. Hartmann-Schröder, 1962b; Fauchald, 1977) the position of the dorsal cirrus on the dorsal notopodial ligule has been used to distinguish these species. In P. gallapagensis View in CoL the dorsal cirrus is attached terminally from anterior chaetigers (from about chaetiger 15), while in P. variegata View in CoL it is terminally attached in posteriormost chaetigers only. Pseudonereis gallapagensis View in CoL can be distinguished from P. atopodon View in CoL by ventral neuropodial ligule as long as acicular ligule in anterior chaetigers compared with up to half as long as in P. atopodon View in CoL , and having fewer paragnaths in Area II than P. atopodon View in CoL ( Table 1).
Fauchald (1977) noted that P. gallapagensis has frequently been confused with other taxa. For this reason the synonymies given above are not extensive but restricted to material examined.
Hartman (1949: 69) examined what she thought to be Kinberg’s specimens of Neanthes variegata , and referred them to P. variegata ( Grube, 1857) , which she obviously considered to be a different species ( Hartman, 1959). This must have been based on a misunderstanding. Kinberg (1865: 172) clearly assigned his specimens to Grube’s species as ‘ N [eanthes] variegata (Grube) ’. Kinberg never described a species under the name Neanthes variegata .
Distribution: Indifatigable Island, Galapagos (type locality); South America, Pacific coast; Hawaii.




| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
| MNHN |
Museum National d'Histoire Naturelle |
| SA |
Museum national d'Histoire Naturelle, Laboratiore de Paleontologie |
| AM |
Australian Museum |
| ZMH |
Zoologisches Museum Hamburg |
| R |
Departamento de Geologia, Universidad de Chile |
| SMNH |
Department of Paleozoology, Swedish Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Pseudonereis
| Bakken, Torkild 2007 |
Pseudonereis rottnestiana
| Hartmann-Schroder G 1979: 118 |
Neanthes cortezi
| de Leon-Gonzalez JA & Solis-Weiss V 2000: 554 |
| Kudenov JD 1979: 120 |
Nereis nichollsi
| Kott P 1951: 95 |
Pseudonereis atopodon
| Chamberlin R 1919: 229 |
Nereis (Pseudonereis) rottnestiana
| Augener H 1913: 187 |
Pseudonereis anomala
| Hylleberg J & Nateewathana A & Bussarawit S 1986: 13 |
| Hutchings PA & Glasby CJ 1985: 108 |
| Hutchings PA & Turvey SP 1982: 141 |
| Gravier C 1901: 197 |
Pseudonereis Kinberg, 1865: 174
| Fauchald K 1977: 90 |
| Kinberg JGH 1865: 174 |
Pseudonereis gallapagensis
| Fauchald K 1977: 32 |
| Hartmann-Schroder G 1962: 432 |
| Hartman O 1949: 68 |
| Kinberg JGH 1865: 174 |
Pseudonereis formosa
| Kinberg JGH 1865: 174 |