
Eidinemacheilus proudlovei, Freyhof, Jörg, Abdullah, Younis Sabir, Ararat, Korsh, Ibrahim, Hamad & Geiger, Matthias F., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4173.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:0E56B22B-B86E-4750-8904-00FD1B2F49B3 |
|
DOI |
https://doi.org/10.5281/zenodo.5693941 |
|
persistent identifier |
https://treatment.plazi.org/id/03E587B3-4F37-721B-21BF-FE5A6CABF978 |
|
treatment provided by |
Plazi |
|
scientific name |
Eidinemacheilus proudlovei |
| status |
sp. nov. |
Eidinemacheilus proudlovei View in CoL , new species
( Figs. 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Holotype. ZFMK-ICH 102318, 53.6 mm SL; Iraq: Sulaimani prov. : Ephemeral spring in the Tabeen Gorge at the slope the northernmost Piramagrun mountain, about 800 m east of the village Kanishok, 35°50'3.40"N, 45° 6'22.14"E; Younis Sabir Abdullah & Korsh Ararat, 31 Mar 2016. GoogleMaps
Paratypes. FSJF 3542, 2 , 45.5, 48.5 mm SL; same data as holotype; Genbank accession number: KX774390 View Materials - KX774392 View Materials .
Diagnosis. Eidinemacheilus proudlovei is distinguished from E. smithi , the only other troglomorphic nemacheilid loach species in the Middle East, and the only other species in Eidinemacheilus known so far, by having 8+8 or 8+7 branched caudal-fin rays (vs. 7+7), no adipose keel on the caudal peduncle (vs. present), enlarged jaws (not enlarged) and a fully developed head canal system (reduced).
Description. See Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 for general appearance and Table 1 View TABLE 1 for morphometric data of holotype and two paratypes. Medium sized, moderately stout species with large head. Body deepest at, or slightly behind nape, profile of back straight, or body depth slightly decreasing towards dorsal-fin origin, and slowly decreasing below dorsal-fin base and towards caudal-fin base. No hump at nape. Greatest body width at pectoral-fin base. Head conical, flattened on ventral surface. Caudal peduncle compressed laterally, 1.8–1.9 times longer than deep. Pectoral fin reaching approximately 60–70% of distance from pectoral-fin origin to pelvic-fin origin. Pelvic axillary lobe absent. Pelvic-fin origin below unbranched or first branched dorsal-fin ray. Pelvic fin not reaching to anus. Anal-fin origin shortly behind anus. Anal-fin origin at vertical of middle between dorsal- and caudal-fin origins. No adipose keel on caudal peduncle. Margin of dorsal fin straight. Caudal fin moderately emarginated. Largest known specimen 53.6 mm SL.
Dorsal fin with 7½ branched rays. Anal fin with 5½ branched rays. Caudal fin with 8+8 (n=1) or 8+7 (n=2) branched rays. Pectoral fin with 10 and pelvic fin with 5 branched rays. Many small unculi on dorsal surface of unbranched and branched pectoral-fin ray 1–3 ( Fig. 5 View FIGURE 5 ). Scales absent. No central, but 1–2 lateral pores on each side in supratemporal canal, 6–7 pores in supraorbital, 7–9 pores in preoperculo-mandibular, 4+8–9 pores in infraorbital canal. Lateral line complete, Reaching to a point above origin. Anterior nostril opening on anterior side of a low, pointed and flap-like tube. Nostrils adjacent to each other, tip of anterior nostril overlaps posterior nostril when folded backwards. No suborbital flap, Groove or slit in fishes examined. Mouth very large, slightly arched ( Fig. 6 View FIGURE 6 ). Lips moderately thick, upper lip smooth or with fine ridges, lower lip with deep furrows. A median interruption in lower lip. Upper lip with median incision or a medial furrow. Lips do not meet medially, leaving mouth parts exposed. Processus dentiformis absent. No median notch in lower jaw. Both jaws large and wide, bordered by a cornified sharp edge. Barbels short, Inner rostral barbell reaching base of outer rostral barbel; outer rostral barbel reaching base of maxillary barbel. Maxillary barbel reaching to base of outer rostral barbel when folded to front. Bony swim bladder capsule ( Fig. 7 View FIGURE 7 ) with large posterior process and very narrow manubrium. A small and narrow epural bone in caudal skeleton.
Coloration. In live, body and fins pink; yellowish white in preservation.
Distribution. Eidinemacheilus proudlovei was washed out from an aquifer into an ephemeral spring close to the village Kanishok. The spring flowed into a small stream, which belongs to the Tabeen drainage. The Tabeen flows to the Little Zab River at the village of Surqawshan. The Little Zab is a tributary of the Tigris. The Tabeen catchment is well known to have many springs and Karezes (underground irrigation systems similar to qanāts in Iran), which are fed by a relatively high winter precipitation in the surrounding mountains.
Etymology. Named for Graham Proudlove (Manchester Museum, the University of Manchester), a world expert on subterranean fishes, for his continuous efforts documenting the diversity of troglomorphic fishes.
Remarks. The genus Eidinemacheilus was recently introduced by Hashemzadeh Segherloo et al. (2016) as a genus-group name for the troglomorphic Nemacheilus smithi , which is endemic to the Tigris drainage in Iran. Eidinemacheilus was distinguished from all genera of nemacheilid loaches in the Middle East by having 7+7 branched caudal-fin rays (vs. 9+8, rarely 8+8), the absence of eyes and pigmentation (vs. presence) and having a reduced set or no head canals (vs. all head canals complete or almost complete). Ideally, the diagnosis of a genus covers all included species and allows to distinguish it from all remaining genera. Nemacheilid loaches are a group of fishes where generic diagnoses are traditionally vague and unclear ( Bănărescu & Nalbant 1995, see also Kottelat 2012), due to a limited set of useful characters as compared to other cypriniform families ( Prokofiev 2009, Prokofiev pers. comm). In his detailed osteological study, Prokofiev (2009) partly failed to recognise the genera of nemacheilid loaches from the Middle East as currently understood, and his results were only partly confirmed by molecular analyses (Freyhof et al. 2014; Freyhof et al. 2015). Hashemzadeh Segherloo et al. (2016) mentioned that all diagnostic characters for Eidinemacheilus are, or could be somehow related to the adaptation of E. smithi to subterranean life. Including E. proudlovei in the same genus renders one character, the head canals (reduced in E. smithi , normally developed in E. proudlovei ) no longer diagnostic for Eidinemacheilus . The low number of branched caudal-fin rays (7+ 7 in E. smithi , 8+7 or 8+ 8 in E. proudlovei ) and the lack of eyes and pigmentation remain diagnostic for Eidinemacheilus , when compared to other nemacheilids in the Middle East.
At the moment, we find it surprising that both subterranean nemacheilid loaches known so far apart within the Tigris drainage belong to the same genus. For now it can only be speculated that both species have evolved from a common (possibly subterranean) ancestor already a substantial time ago — several million years, if we assume a molecular clock-like behaviour of COI. Given that we only studied one mitochondrial marker (COI) we did not opt to perform an in-depth analysis of the absolute divergence time. In addition, there are no robust calibration points, such as geologic events, and rates of molecular evolution are expected to be heterogeneous among different lineages (e.g. Lavinia et al. 2016; Wilke et al. 2009), a problem especially given our heterogeneous sample composition. Yet (and despite the uncertainties of mitochondrial DNA molecular clock calibration) the highly divergent COI clusters (>8% K2P-distance) of the two Eidinemacheilus species are indicative of at least four million years of unique evolutionary history, using the rate proposed in several studies ( April et al. 2011; Brown et al. 1979; Durand et al. 2003; Zardoya & Doadrio 1999).
The circumstances of the discovery of E. proudlovei , and the never before reported wash-out of fishes from this aquifer, raise the question how many other populations and species of subterranean fishes might still exist unearthed in the Middle East and elsewhere.
Comparative material. Eidinemacheilus smithi : FSJF 3431, 1 , 28 mm SL; FSJF 3541, 4 , 30–38 mm SL; Iran: Lorestan prov.: Natural well-like opening being the origin of the stream Kay-e-Ru flowing to Sirom River, near Baq-e-Levan oasis, 33°04'38''N 48°35'35''E. GoogleMaps




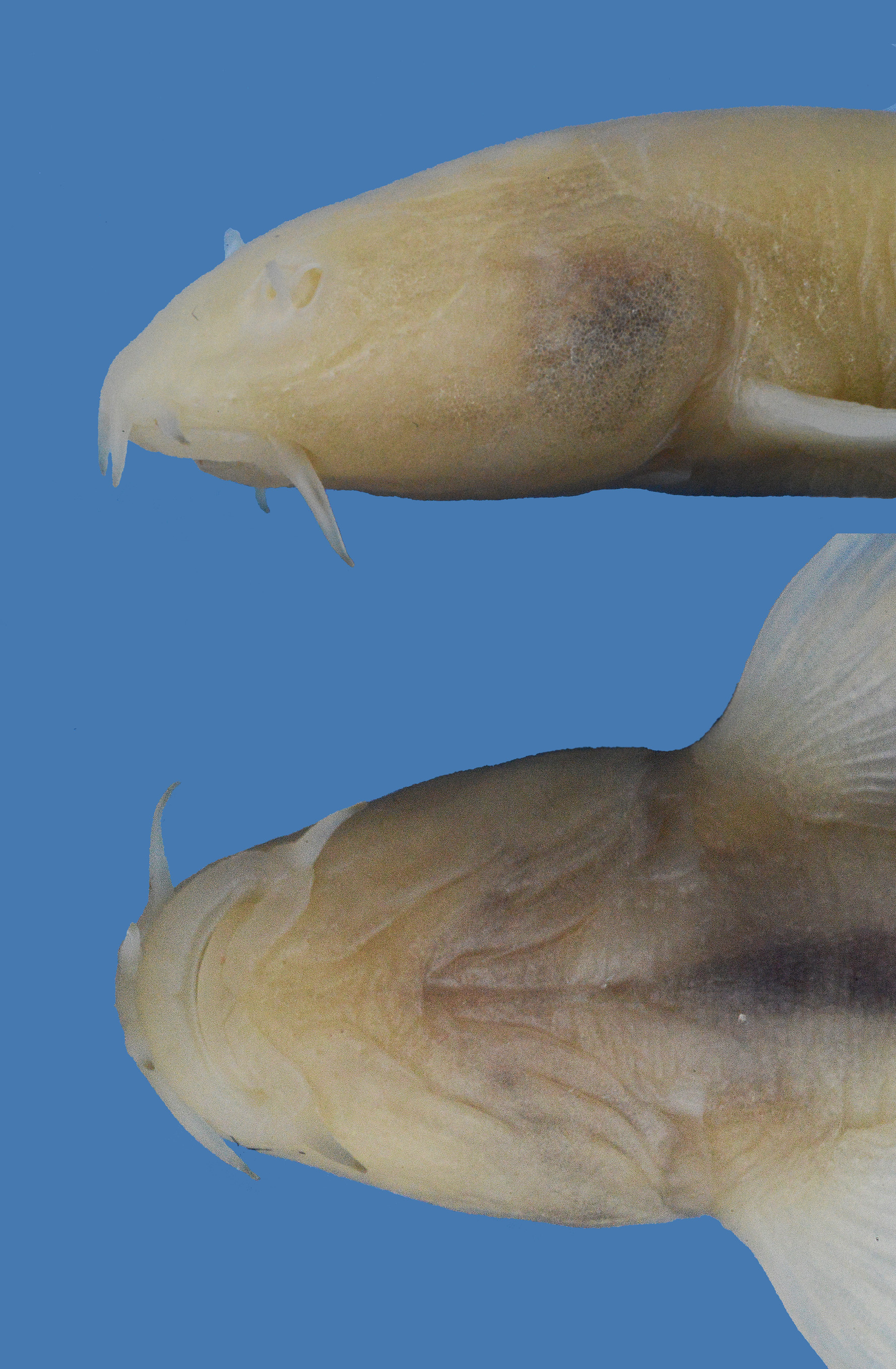
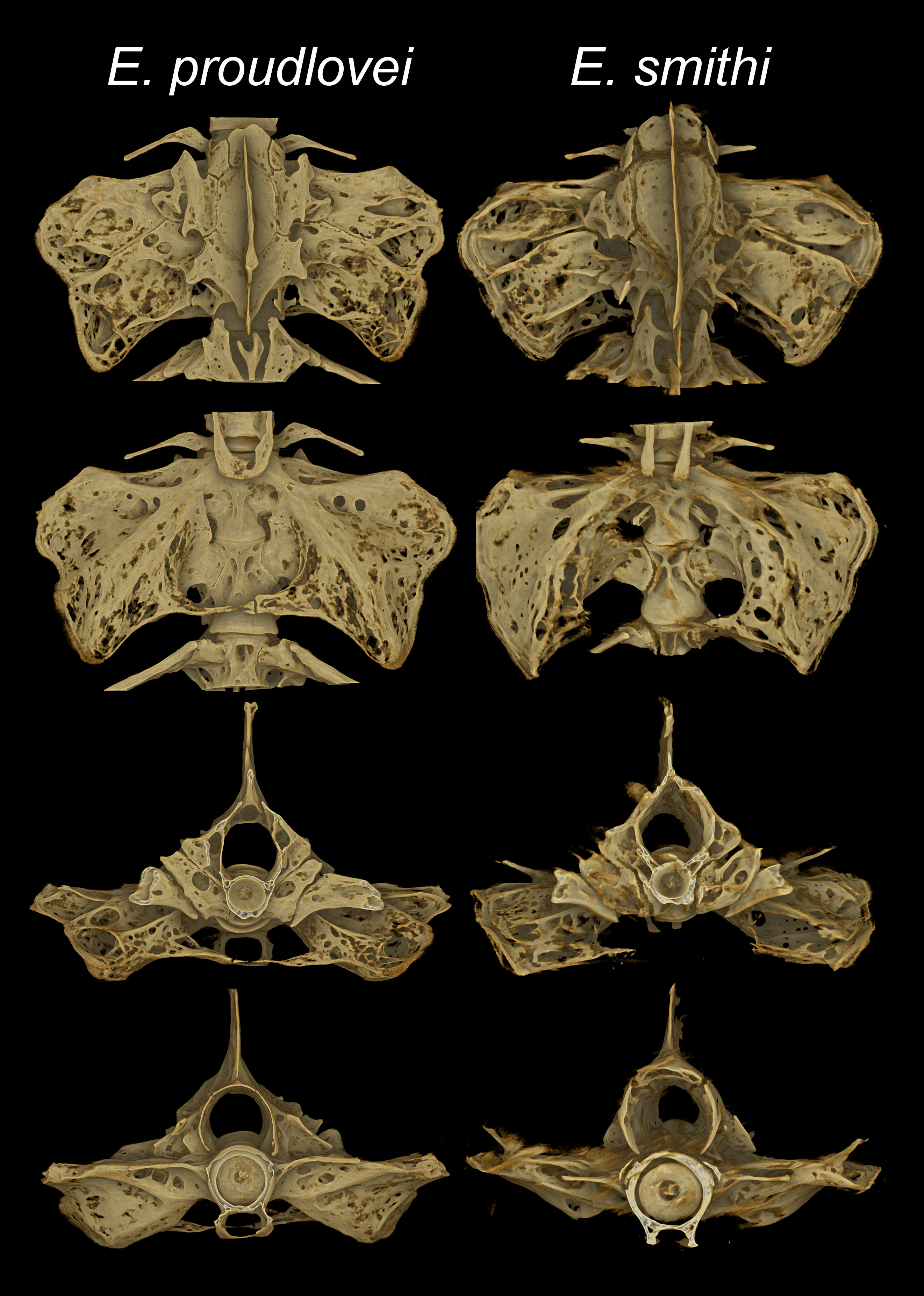
TABLE 1. Morphometric data of Eidinemacheilus proudlovei (ZFMK-ICH 102318, holotype; FSJF 3542; paratypes; n = 3).
| holotype | paratype | paratype | |
|---|---|---|---|
| SL (mm) | 53.6 | 45.5 | 48.5 |
| In percent of standard length | |||
| Head length | 28.9 | 26.4 | 27.0 |
| Body depth at dorsal-fin origin | 14.4 | 15.6 | 17.1 |
| Predorsal length | 57.0 | 35.5 | 57.0 |
| Postdorsal length | 34.0 | 34.7 | 36.6 |
| Prepelvic length | 55.8 | 57.2 | 56.8 |
| Preanal length | 75.7 | 77.8 | 78.0 |
| Distance between pectoral and pelvic-fin origins | 30.2 | 31.7 | 32.7 |
| Distance between pelvic and anal-fin origins | 20.3 | 20.9 | 19.9 |
| Distance between anus and anal-fin origin | 2.7 | 2.5 | 2.5 |
| Caudal-peduncle depth | 8.3 | 9.3 | 9.4 |
| Caudal-peduncle length | 15.8 | 18.0 | 17.2 |
| Dorsal fin-base length | 10.3 | 9.0 | 10.3 |
| Anal fin-base length | 8.4 | 7.8 | 8.4 |
| Pelvic-fin length | 12.1 | 10.7 | 11.8 |
| Pectoral-fin length | 18.4 | 18.3 | 18.8 |
| In percent of head length | |||
| Head depth at nape | 14 | 14 | 15 |
| Maximum head width | 18 | 16 | 18 |
| Mouth width | 8 | 8 | 8 |
| Inner rostral-barbel length | 4 | 4 | 4 |
| Outer rostral-barbel length | 7 | 6 | 5 |
| Maxillary-barbel length | 5 | 5 | 4 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |