
Simulium tenebrosum
|
publication ID |
https://doi.org/10.1093/isd/ixz004 |
|
persistent identifier |
https://treatment.plazi.org/id/03E187EF-FFB6-AE49-FF09-B84E1D5B1CF1 |
|
treatment provided by |
Felipe |
|
scientific name |
Simulium tenebrosum |
| status |
|
The banding patterns of all 107 chromosomally prepared larvae were analyzed completely, including two males infected with a microsporidium resembling Amblyospora varians (Léger) sensu Vávra and Undeen (1981) (Microsporidia: Amblyosporidae ) and
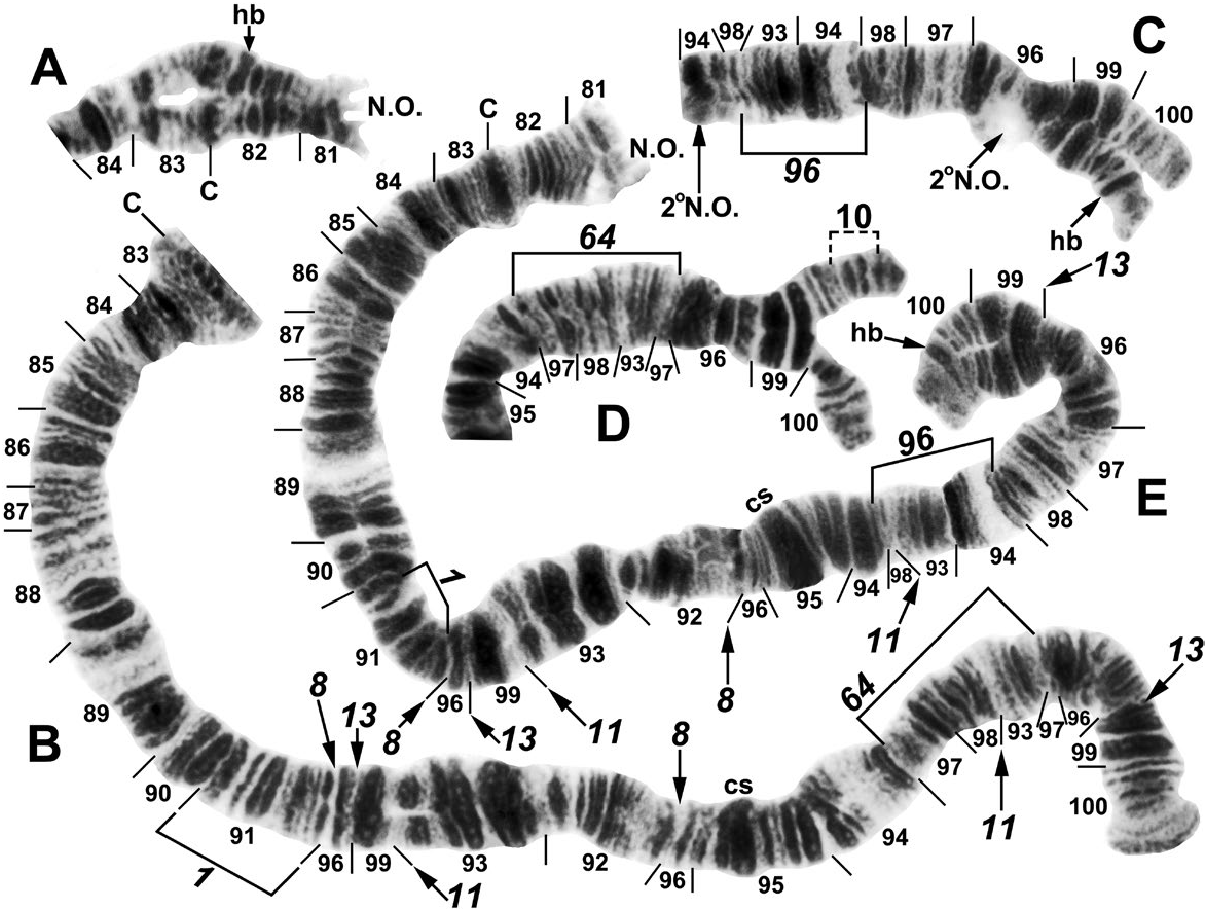
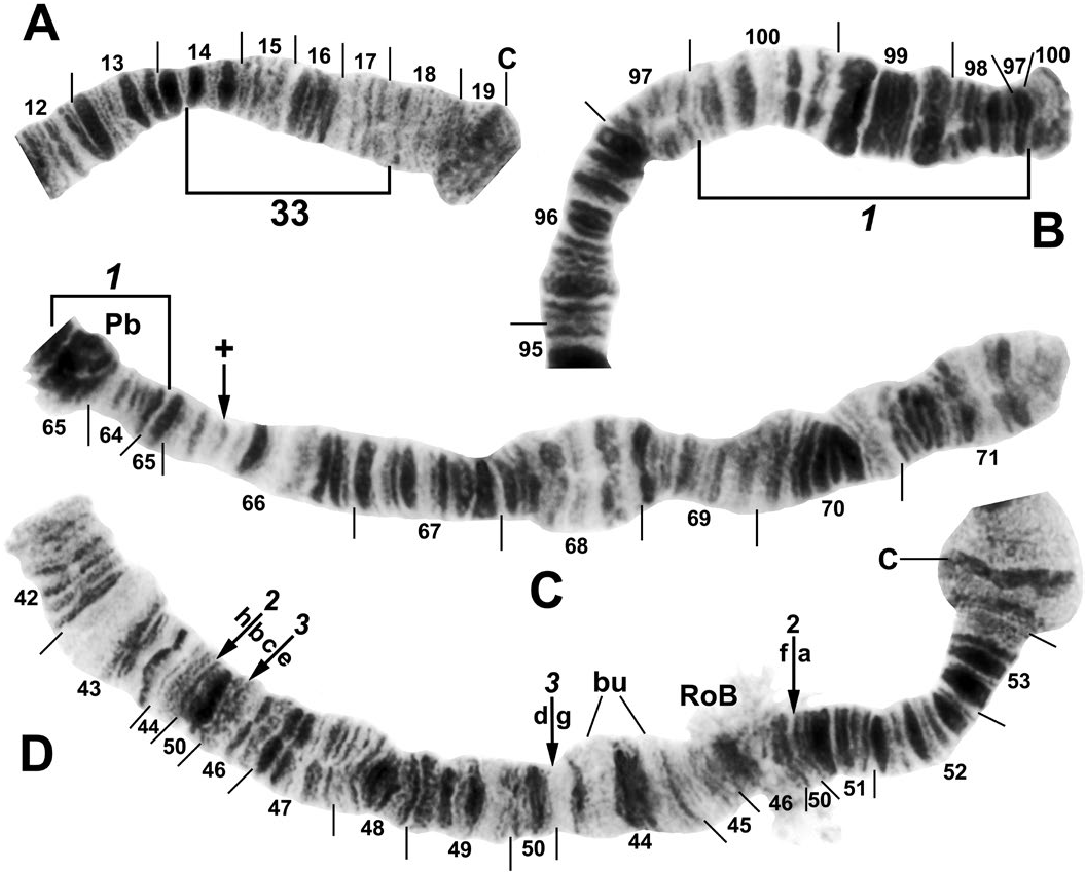
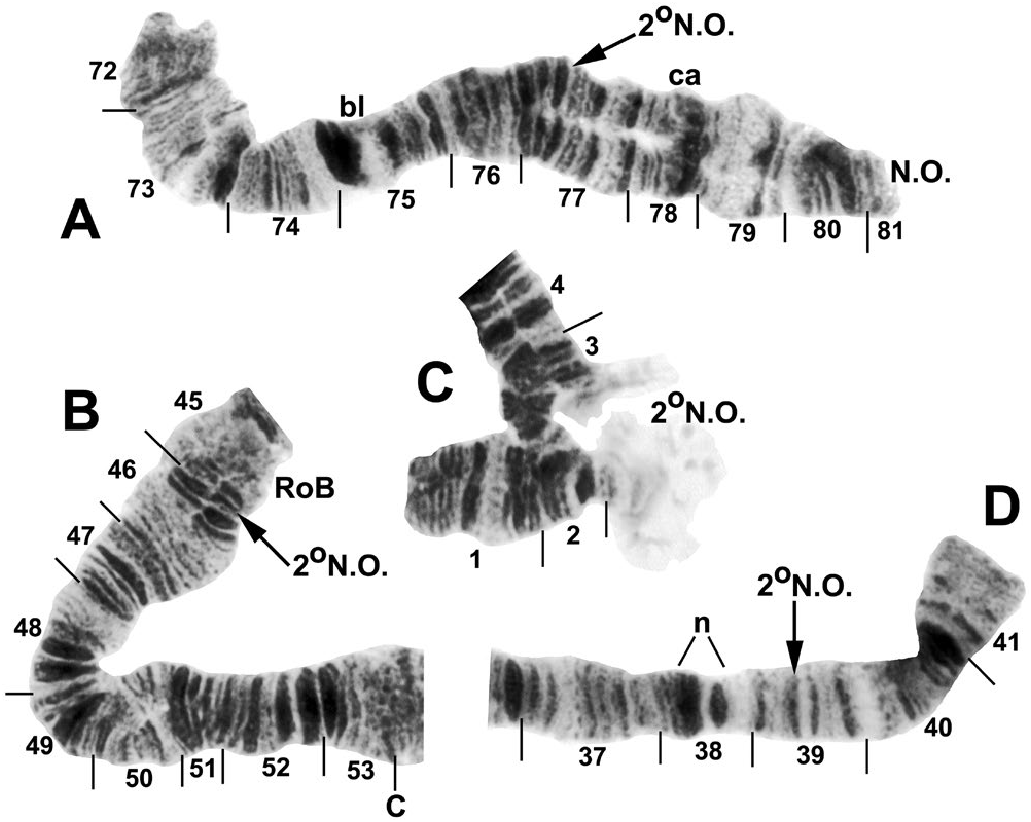
one female infected with an unidentified mermithid nematode ( Mermithidae ). Homologs were tightly paired (98–100%). All larvae had the standard chromosomal banding sequence for the Southeast Asian S. tuberosum lineage (sensu Adler et al. 2016c) and were also fixed for IIIL-8, IIIL-11, and IIIL-13 ( Fig. 3 View Fig ). All larvae additionally had either of two mutually exclusive fixed-inversion sequences in the IIIL arm: IIIL-64 or IIIL-96 ( Fig. 3 View Fig ). The complete absence of individuals heterozygous for either of these inversions indicated the presence of two separate breeding populations (i.e., species): 57.9% ( 35 females and 27 males) with IIIL-64 and 42.1% ( 24 females and 21 males) with IIIL-96 ( Table 2). A presorted subsample of 33 final-instar larvae yielded 82% correct assignment to one or the other chromosomal sequences, with IIIL-64 representing the largest larvae (94% correct, n = 18) and IIIL-96 the smallest (67% correct, n = 15). Larvae of intermediate size carried either IIIL-64 or IIIL-96, indicating considerable size overlap ; only the extreme sizes agreed reasonably well with the two chromosomal sequences.
A small subterminal inversion, IIIL -10 ( Fig. 3D View Fig ), was distributed nonrandomly among larvae ( Table 3). Most of the 27 males (90.9%) with the IIIL-64 sequence carried IIIL -10 heterozygously. The remaining 9.1% of males with IIIL-64, plus all males with IIIL-96 and all females, lacked the inversion. IIIL -10, therefore, marked a differentiated Y chromosome (Y 1) in males with IIIL-64 ; the remainder (9.1%) of the males with IIIL-64 were, thus, Y 0. IIIL- 64 females ( n = 35) were either standard (X 0, 77.1%) for section 100 of IIIL or had heteroband 100hb1 (X 1, 22.9%). Two males were heterozygous for IIIL -10 and IIIL 100 hb1, but on different homologs, and were, therefore, X 1 Y 1. All individuals with IIIL-96 had cytologically undifferentiated sex chromosomes (X 0 Y 0) .
Autosomal polymorphisms were limited to one inversion (IS-33, Fig. 2A View Fig ), six different secondary nucleolar organizers ( Figs. 3C View Fig and 4 View Fig ), three of which were in a single female larva, and two different heterobands ( Fig. 3A, C, E View Fig ); all polymorphisms had frequencies less than or equal to 0.02 ( Table 2). Heteroband 100hb1, which was X-linked in individuals with IIIL-64, was expressed heterozygously as an autosomal polymorphism ( Fig. 3C and E View Fig ) in two female larvae with IIIL-96 at an overall frequency of 0.02.
We assigned informal handles to the two groups of S. tenebrosum : cytospecies A for individuals fixed for IIIL-64 and cytospecies B for individuals fixed for IIIL-96.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |