
Xanthopan praedicta Rothschild & Jordan, 1903
|
publication ID |
https://doi.org/10.5281/zenodo.5856141 |
|
DOI |
https://doi.org/10.5281/zenodo.5856146 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF87EC-FFD8-8D52-CC96-C375A755BA4A |
|
treatment provided by |
Donat |
|
scientific name |
Xanthopan praedicta Rothschild & Jordan, 1903 |
| status |
|
Morphological differences between Xanthopan m. morganii and X. m. praedicta
Illustrations in ARDITTI & al. (2012) may give the impression that “ Xanthopan morganii morganii ” is a highly variable taxon, but some of the specimens pictured were erroneously referred as originating “from Gabon” ( ibid., fig. 7 View FIG A-C) when actually being from Madagascar, and thus representing “ Xanthopan morganii praedicta ”.
The main differences between X. m. morganii and X. m. praedicta are listed hereafter, following an anteroposterior order for the enumeration of the corresponding characters.
Head.
1, proximal section of the proboscis – morganii : reddish brown; praedicta : black.
2, ventral vestiture of segment 1 of the labial palpus (in lateral view) – morganii : with numerous dark grey scales, at least distally, so that the coloration of segment 1 contrasts moderately with the brown region of segment 2; praedicta : relatively light-coloured, contrasting strongly with the dark brown region of segment 2.
Foreleg.
3, tarsus in dorsal view – morganii : basitarsus lightcoloured, contrasting strongly with tarsomeres 2-5, which are black or blackish (with some white at the extremities of tarsomere 2); praedicta : tarsomere 2 at least partially brown, with an intermediate coloration between that of the basitarsus (ochre or pale brown) and that of tarsomeres 3-5 (dark brown).
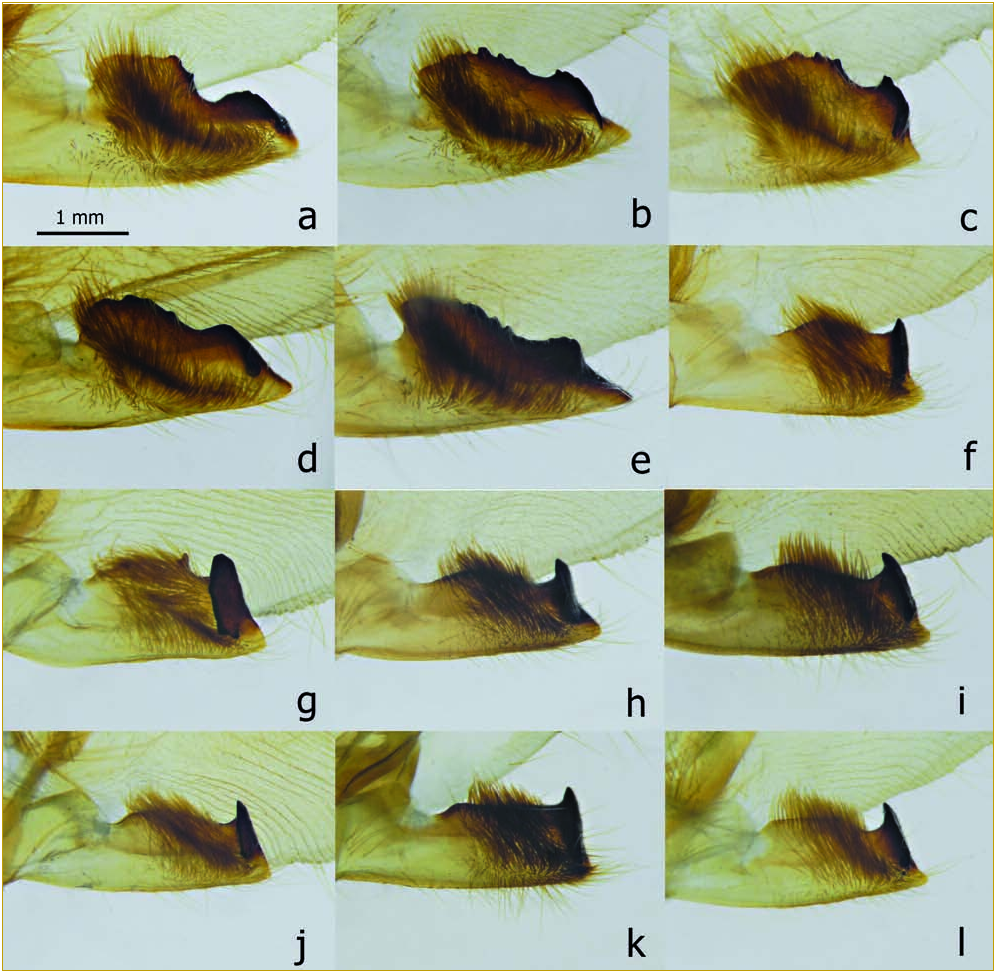
Forewing shape.
4, termen – morganii : angulate at M 3 (fig. 2 View FIG , a-d); praedicta : regularly arched between CuA 2 and the apex (fig. 2 View FIG , e-h).
Forewing (dorsal surface).
5, dark subbasal costal mark – morganii : a trian - gular spot, extending distad along the costa (fig. 2 View FIG , a-c); praedicta : not distinctly triangular, usually in the form of a short zigzag, which is roughly parallel to vein CuA 2 and often unites two very small dark spots (the costal spot being neither clearly triangular nor markedly prolonged distad along the costa: fig. 2 View FIG , e and g).
6, lower discal light-coloured spot (on discocellular m2-m3) – morganii : typically broad, greyish white, without well defined dark edges, and often slightly extended on the base of vein M 2 (fig. 2 View FIG , a-c); praedicta : thin, circumflexshaped, white or whitish, relatively well delimited, and not extended distad on M 2 (fig. 2 View FIG , e and g).
7, region between veins M 1 and M 3 – morganii : greyish or beige (fig. 2 View FIG , a-c), sometimes (proximally) with a narrow pinkish band between M 2 and M 3 (fig. 2 View FIG , c); praedicta : typically, with a large chestnut-brown area, which is ill-delimited above M 2 and does not extend beyond the (often obsolete) submarginal line (fig. 2 View FIG , e and g).
8, blackish subapical streak below vein Rs 4 – morganii : usually thin (fig. 2 View FIG , a-c), though sometimes fairly broad (plate IV, fig. 7 View FIG , in CARCASSON, 1976); praedicta : usually broader (fig. 2 View FIG , e and g; contrary to a statement in ROTHSCHILD & JORDAN (1903), the broad streak is not a constant characteristic of the taxon praedicta ).
Forewing (ventral surface).
9, apical region – morganii : uniformly brownish (fig. 2 View FIG , d); praedicta : with an upper light-coloured area that contrasts with the lower – brown – apical area (fig. 2 View FIG , f and h; the light-coloured area is occasionally faintly indicated, notably in “faded” specimens; it was found to be almost absent in a specimen in fairly good condition collected by one of us (PB) in Lakato forest, near Moramanga).
Hindwing (dorsal surface).
10, central and anal light-coloured patches – morganii : orange yellow, contrasting with the whitish colour of the discal cell and costal region (fig. 2 View FIG , a-c); praedicta : relatively pale yellow, having approximately the same colour as the cell and the costal region (fig. 2 View FIG , e and g).
11, yellow central patch – morganii : well-developed and distinctly triangular (fig. 2 View FIG , a-c); praedicta : usually a bit smaller and subtriangular (fig. 2 View FIG , e and g).
12, dark central region – morganii : uniformly brown (fig. 2 View FIG , a-c); praedicta : usually greyish brown, with dark brown longitudinal stripes between the veins (fig. 2, g).
13, dark streak of the anal angle – morganii : usually well indicated, edged with whitish or pale grey (fig. 2 View FIG , a-c); praedicta : usually less conspicuous (fig. 2 View FIG , e and g).
14, fringe of the termen – morganii : dark brown at vein apices, whitish or greyish between veins; praedicta : pale chestnut (at vein apices) and yellowish (between veins).
Hindwing (ventral surface).
15, light-coloured patch below the discal cell – morganii : yellow, well indicated, extending from wing base to the median section of CuA 2 and the basal region of CuA 1 (fig. 2 View FIG , d); praedicta : rather inconspicuous and ill-delimited, never large, usually yellowish or pinkish (fig. 2 View FIG , f and h).
Thorax and abdomen (coloration).
16, ventral region – morganii : white or yellowish white (fig. 2 View FIG , d); praedicta : always pink (fig. 2 View FIG , f and h), as already mentioned in the original description ( ROTHSCHILD & JORDAN, 1903: 32).
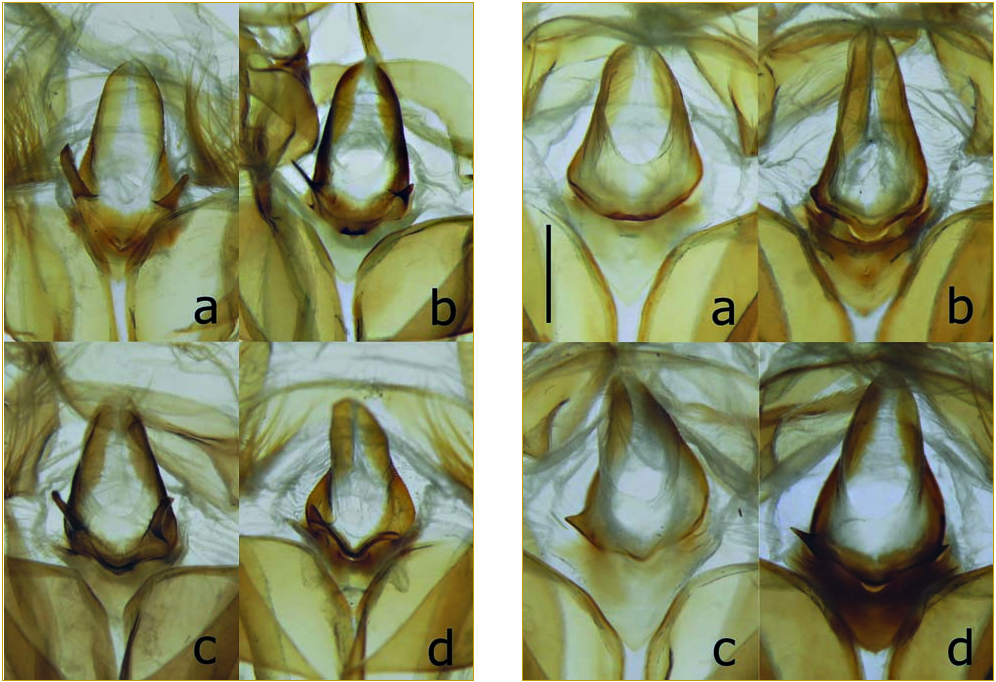
Male genitalia.
17, in the phallus, ratio between a (= distance between the anterior “elbow” and the posterior extremity of the bulbus ejaculatorius) and b (= length of the sclerotized phallus) – morganii : a long, varying from 48 to 57 percent of b (fig. 3 View FIG , b); praedicta : a usually shorter, varying from 41 to 48 percent of b (fig. 3 View FIG , f and h).
18, saccus – morganii : elongate (fig. 3 View FIG , a and c; fig. 7, g and h); praedicta : shorter and more robust (fig. 3 View FIG , e and g; fig. 7 View FIG , e and f).
19, valva – morganii : relatively narrow, its greatest breadth (c) being less than 1/3 of its total length (d) (fig. 3 View FIG , a and c); praedicta : broader, c being more than 1/3 of d (fig. 3 View FIG , e and g). These distances have been measured on spread genitalia, with c extending transversely from the most convex part of the costa to the ventral edge of the valva, below the praesacculus.
20, praesacculus (“harpe”) – morganii : usually with smooth dorsal edge (disregarding pilosity), except for a strong, undivided, dorso-distal tooth (fig. 3 View FIG , a and c; fig. 4 View FIG , f and h-l), occasionally with a supernumerary small tooth proximad of the dorso-distal tooth (fig. 4 View FIG , g); praedicta : dorsal edge of the praesacculus irregular, with several teeth proximad of the broad dorso-distal projection, which is not very prominent and usually has a crenulate edge (fig. 3 View FIG , e and g; fig. 4 View FIG , a-e). According to figure 53 in GRIVEAUD (1959), the dorso-distal projection of the praesacculus of X. praedicta may occasionally resemble that of X. morganii but it should be noted that his figure is neither a photograph nor a detailed drawing. KITCHING (2002: 489, character 65) had already noticed the difference between X. morganii and X. [“ m.”] praedicta with regard to the dorsal edge of the proximal region (“lobe”) of the “harpe”, described as “rather” truncate and smooth in the case of the former taxon, provided with “several blunt, irregular teeth” in the case of the latter.
21, juxta – morganii : exhibiting on the left side a small lateroventral projection, which is usually elongate (fig. 3 View FIG , a and c; fig. 5 View FIG , a-c), although occasionally short and triangular (as in a specimen from Grande Comore: fig. 5 View FIG , d); praedicta : without well-developed lateroventral projection on the left side (fig. 3 View FIG , e and g; fig. 6 View FIG , a and b) or, at most, with a short, triangular lateroventral projection (fig. 6 View FIG , c and d).
22, apex of the gnathos (observed ventrally between slide and coverslip) – morganii : distinctly notched mesally (fig. 3 View FIG , a and c; fig. 7 View FIG , c and d); praedicta : blunt or convex mesally, never notched (fig. 3 View FIG , e and g; fig. 7 View FIG , a and b).
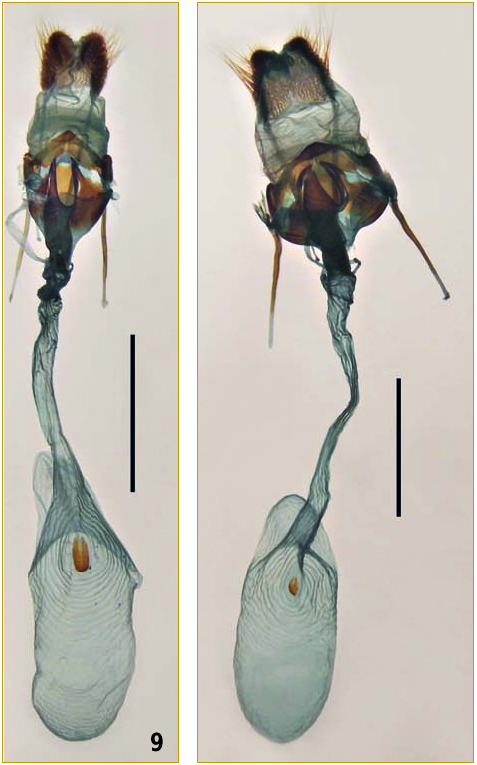
Female genitalia.
23, posterior pouch of the corpus bursae (delimited ventrally, in lateral view, by the anterior, widened region of the ductus bursae) – morganii : fairly deep (fig. 10 View FIG , c-e), its depth (x) being greater than or equal to one fifth of the total length (y) of the corpus bursae; praedicta : apparently less deep (fig. 10 View FIG , a and b), its depth (x) being less than one fifth of y. This difference in the shape of the corpus bursae is more difficult to appreciate when the genitalia are observed in ventral view (figs. 8 View FIGS and 9 View FIGS ).
24, signum – morganii : well developed, elliptical and more or less horseshoe-shaped (fig. 11 View FIG , d-h); praedicta : smaller (fig. 11 View FIG , a and b) to almost absent (fig. 11 View FIG , c).
25, ostium bursae and sterigma – morganii : lateral sclerotized plates of the lamella antevaginalis posteriorly fused with the lateral edges of the ostium bursae (fig. 12 View FIG , d-f; also fig. 13 View FIG B in KITCHING, 2002) so that the plates and the ostium form a single pair of posterior projections in lateral view (fig. 12 View FIG , j-l); praedicta : posterior extremities of the lateral plates of the lamella antevaginalis not merged with the posterior extremities of the edges of the ostium bursae (fig. 12 View FIG , a-c) so that the plates and the ostium form two pairs of posterior projections in lateral view (fig. 12 View FIG , g-i).
In the above list, at least seven morphological characters provide clear-cut differences between the two taxa (viz. numbers 3, 4, 10, 16, 22, 25, and also 20 if one takes into account only the proximal region of the praesacculus). The differences corresponding to characters 1, 2, 5, 14, 15, and 18 may also be clear-cut ones, although some of them (such as 14) would need to be examined on fresh specimens (rather than on collection specimens, which often tend to be faded). By contrast, numbers 4, 17, 19 and 23 correspond to insufficiently studied quantitative differences, and numbers 6, 7, 8, 9, 11, 12, 13, and 21 provide differences that are observable in most, though not all, cases.
On account of (1) a deep, ancient genetic divergence (dated to ca. 7.4 Mya by NETZ & RENNER, 2017) and (2) the above-mentioned clear-cut morphological differences (e. g. in the ostium bursae), Xanthopan morganii (Walker, 1856) and Xanthopan praedicta Rothschild & Jordan, 1903 stat. nov. should henceforth be regarded as distinct species (no longer as “subspecies”), the latter being endemic to Madagascar while the former inhabits sub-Saharan Africa and the Comoro Islands.
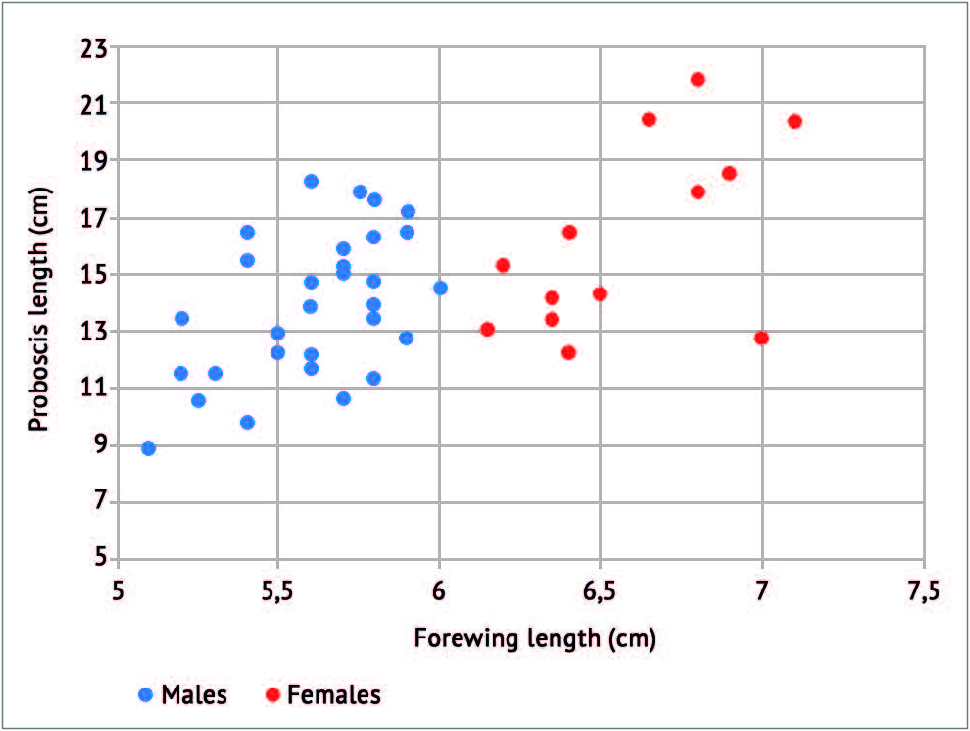
As emphasized in the next paragraph, the proboscis and forewing lengths are usually greater in Xanthopan praedicta than in Xanthopan morganii (e. g. fig. 13 View FIG ) but they vary in both species, with overlap zones in the measurements.
Proboscis length variability in the two species of Xanthopan
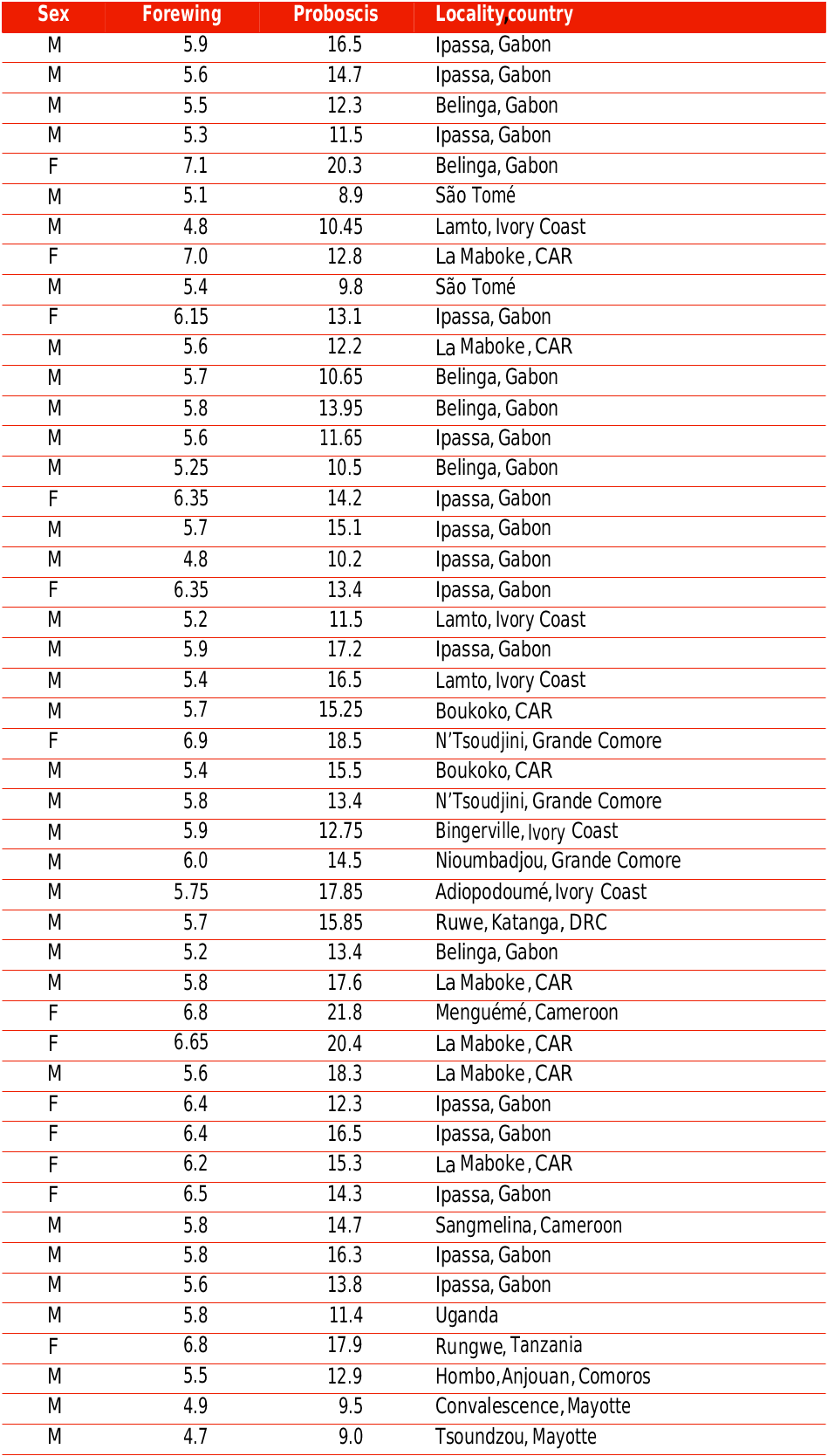
Although the first measure of the proboscis length (22.5 cm) of Xanthopan praedicta was given in the original description of this taxon ( ROTHSCHILD & JORDAN, 1903), very few studies have been devoted, up to now, to the precise knowledge of this length, with assessment of its variability ( ARDITTI & al., 2012: Table 4). Such data are even scarcer in scientific literature in the case of Xanthopan morganii . For the latter, the length of the proboscis would be “seven inches and a half” (19 cm) according to WALLACE (1891: 148), 12 cm according to MARTINS & JOHNSON (2007; Table 3 – one male), and would have an average value of either 13.8 cm ( RYCKEWAERT & al., 2011 – 6 individuals) or ca. 15 cm ( BALDUCCI & al., 2019 – mean based on 12 individuals). In fact, after measuring the proboscides of 47 specimens from the MNHN (Additional data, Table 1 View Additional ), we find an average length of 14.18 cm for X. morganii , with the range being 8.9-21.8 cm (it should be noted that the mention “over 22 cm” in PINHEY (1962) concerns the taxon praedicta , not the true morganii ; Table 4 of ARDITTI & al. (2012) may be misleading on that point). We managed to measure a higher number of proboscides in the case of X. praedicta (Additional data, Table 2 View Additional : 77 individuals, most from MNHN and NHMUK collections, 24 released after measurements (Ranomafana National Park, DCL, March 2020)): we thus have for the proboscis of this Malagasy hawkmoth an average length of 20.8 cm – higher than the means proposed by NILSSON & al. (1985) and RYCKEWAERT & al. (2011), namely, respectively, 19.6 cm (from 8 individuals) and 18.26 cm (from 5 individuals) – with the range being 15.2-28.5 cm (nevertheless RYCKEWAERT & al. found a shorter proboscis (14.5 cm long) in an individual of the Malagasy Xanthopan ).
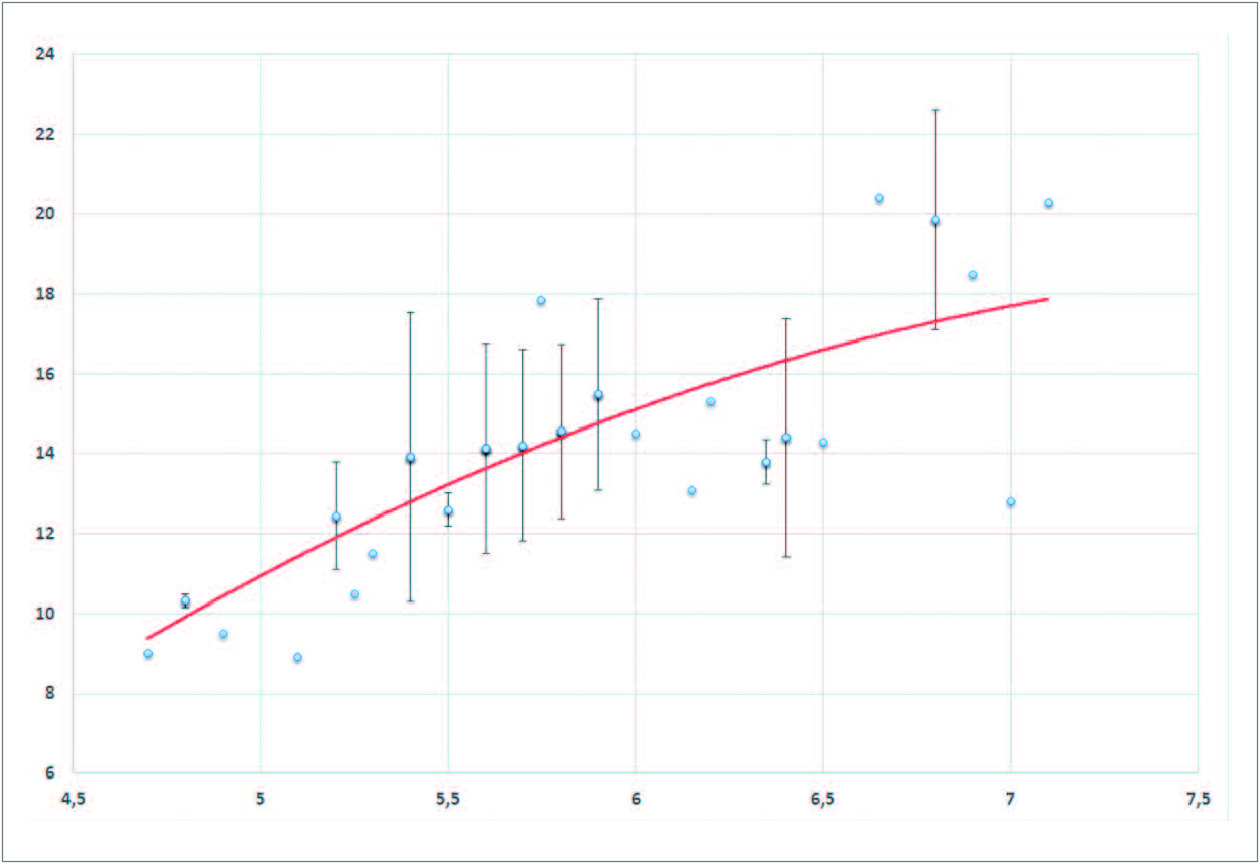
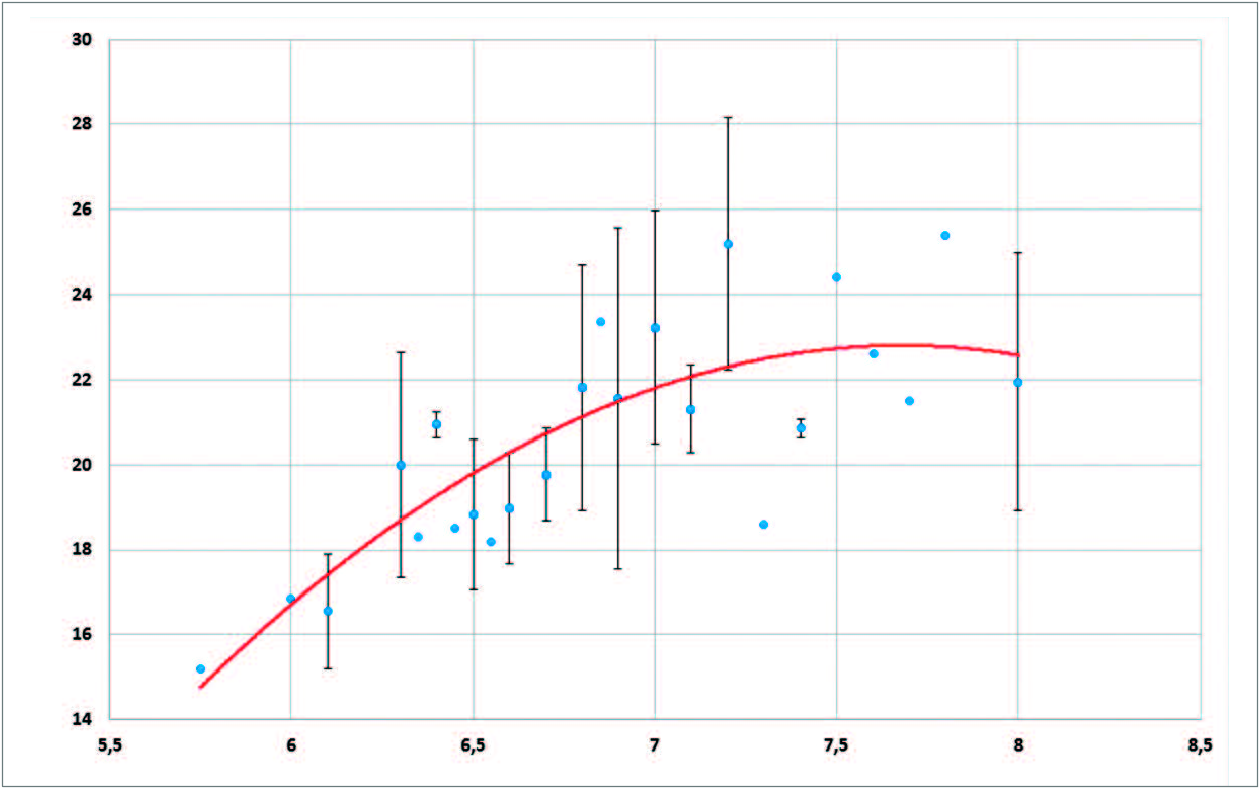
While the proboscis lengths of X. morganii and X. praedicta overlap partially (fig. 14 View FIG ), the commonest lengths are still more remote from one another (12-13 cm; 20 cm) than are their respective means (ca. 14.2 cm; 20.8 cm). Examining the length of the proboscis in terms of moth size (appraised via forewing length) and taking into account the sexes (fig. 15 View FIG ), females of X. morganii are routinely larger than the males of this taxon and usually have longer proboscides (16.2 cm on average; range: 12.3-21.8 cm – contra 13.4 cm (range: 8.9-18.3 cm) in the males), with a few values over 20 cm (three in our dataset). A 2 nd order polynomial trendline made with Excel (fig. 17 View FIG ) shows that proboscis length is roughly proportional to forewing length in the case of X. morganii . By contrast, in X. praedicta , the longest proboscides were found in males (fig. 16 based on the data in Additional data, Table 2 View Additional ), with one value (28.5 cm, from Ranomafana) seemingly representing an absolute record for a lepidopteran proboscis length (28 cm, in certain individuals of Amphimoea walkeri (Boisduval, [1875]) , had been previously regarded as the highest figure among insect haustellum lengths: MILLER, 1997). Nevertheless, in X. praedicta too, the average length of the proboscis is higher in females (21.97 cm; range: 18.6-25.4 cm) than in males (20.65 cm; range: 15.2-28.5 cm). For this species, the 2 nd order polynomial trendline ( fig. 18 View FIG ) is distinctly curved and becomes more or less horizontal for the larger specimens (females), which suggests that proboscides significantly exceeding 28.5 cm are unlikely to be found in Madagascar.
Discussion and conclusion
Despite the absence of an absolute difference in proboscis length, the proboscis of Xanthopan praedicta is usually much longer than that of X. morganii , which assuredly reflects adaptations to different sets of visited flowers, especially among long-spurred orchids. Since most sphingophilous flowers are visited at night, their pollinators remain poorly known, notably in tropical regions. To date, only three African orchids have been recorded as sphingophilous flowers visited or pollinated by X. morganii , namely Aerangis kotschyana , Rangaeris amaniensis , and Bonatea steudneri ( MARTINS & JOHNSON, 2007; BALDUCCI & al., 2019). None of these three Orchidaceae occurs in Madagascar, an island which nevertheless houses numerous species of long-spurred orchids ( BOSSER & LECOUFLE, 2011; NETZ & RENNER, 2017). The predicted pollination of Darwin’s Comet Orchid ( Angraecum sesquipedale , endemic to Madagascar) by X. praedicta was eventually confirmed by a viscidium found on a moth in 1992 and photographs of it visiting flowers of the orchid (see ARDITTI & al., 2012: 419), also thanks to behavioural observations in captivity ( WASSERTHAL, 1993, 1996, 1997) and, most recently, in the wild, with a film (made by Philip DEVRIES in 2004) that can still be watched on the Web (https://www.youtube.com/watch?v=OMVN1EWxfAU). However X. praedicta may possibly not be the sole pollinator of A. sesquipedale , even though this orchid has an exceptionally long nectar tube (the “spur” ranging from 27 to 43 cm according to WASSERTHAL, 1997: 344). Indeed Agrius convolvuli (Linnaeus, 1758) was observed visiting flowers of A. sesquipedale in a large flight tent, although without removing the pollinaria of these flowers ( WASSERTHAL, 1997). It should be noted that, with an average length of 11 cm (range: ca. 8.5-15.5 cm – RYCKEWAERT & al., 2011), the proboscides of Malagasy specimens of Agrius convolvuli are markedly longer than those of conspecifics from Europe or mainland Africa ( NILSSON & al., 1985: 8). Moreover, although – quite surprisingly – reported only recently (e. g. NILSSON & al., 1985), Madagascar possesses another Sphinginae equipped with a very long proboscis, namely Coelonia solani (Boisduval, 1833) , which has an average tongue length of 17.6 cm (range: 13-24 cm according to RYCKEWAERT & al., 2011). Although C. solani is the main pollinator of Angraecum sororium ( WASSERTHAL, 1997) , the sister species of A. sesquipedale ( NETZ & RENNER, 2017) , we cannot completely rule out a possible role of this hawkmoth in the pollination of Angraecum sesquipedale (despite the fact that it did not visit two flowers of this orchid in a greenhouse in Germany: ibid., p. 353). On the other hand, X. praedicta is known to visit flowers of different species, some in Orchidaceae ( Angraecum sesquipedale and Angraecum compactum ; perhaps also Aerangis fuscata: NILSSON & al., 1987 ), some in other families: Malvaceae (the baobab Adansonia perrieri: BAUM, 1995 ), Lamiaceae ( Clerodendrum: WASSERTHAL, 1997), Verbenaceae ( Lantana), Amaryllidaceae ( Crinum), and probably several others. In fact, there are complex interactions between orchids and their pollinators ( HOULIHAN & al., 2019), so that we can rule out KRITSKY’ s (1991) hypothesis that there would be in Madagascar “a gigantic moth with a proboscis even longer” than that of X. praedicta in view of the existence of Angraecum longicalcar , an orchid with a nectary “nearly 40 cm long, 10 cm longer than that of A. sesquipedale ”. It should be noted that, actually, the “spur” of A. sesquipedale ranges from 27 to 43 cm and contains a varying volume of nectar, a parameter that cannot be neglected in studies about pollination ( WASSERTHAL, 1997: Fig. 4 View FIG ). According to NETZ & RENNER (2017), the shape of the labellum should also be taken into account: it may form a landing platform for large hawkmoths, as in A. sesquipedale , or not, as in A. longicalcar (their Fig. 1B shows Xanthopan praedicta “trying to visit” flowers of this rare orchid). It should be noted that botanists from Royal Botanic Gardens, Kew, using camera traps, failed to observe any pollinator, not even X. praedicta , at the sole known wild population of A. longicalcar (https://www.theguardian.com/world/ 2017/may/13/madagascar-mass-extinction-plants-kewgardens).
The taxonomic change we propose here not only for the first time gives appropriate weight to clear-cut morphological differences (including in female genitalia) and high mitochondrial divergence. It also gives long deserved recognition, at the species level, to one of the most celebrated of all Malagasy endemics.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sphinginae |
|
Genus |