
Oedichirus geniculatus (Sahlberg)
|
publication ID |
https://doi.org/10.1206/816.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF8794-7D48-D130-FD57-5641FCCA07C8 |
|
treatment provided by |
Felipe |
|
scientific name |
Oedichirus geniculatus (Sahlberg) |
| status |
|
Oedichirus geniculatus (Sahlberg) View in CoL
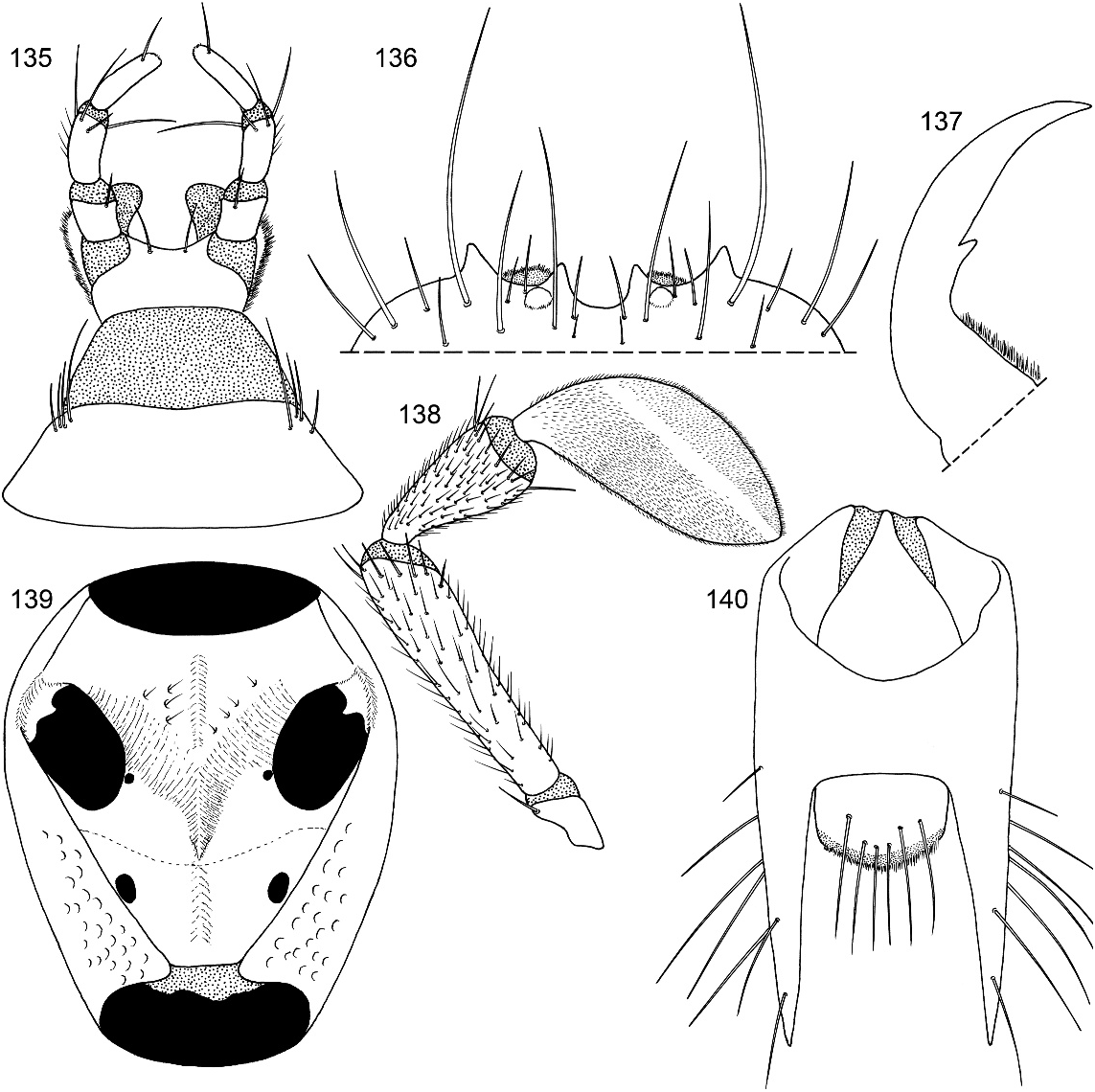
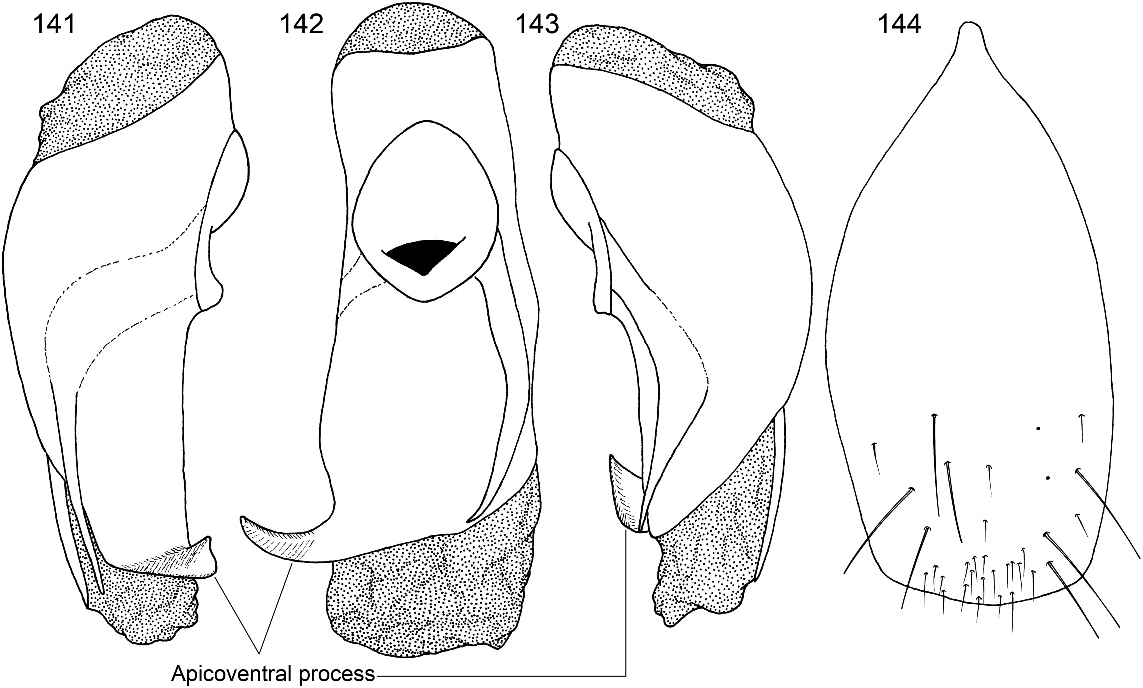
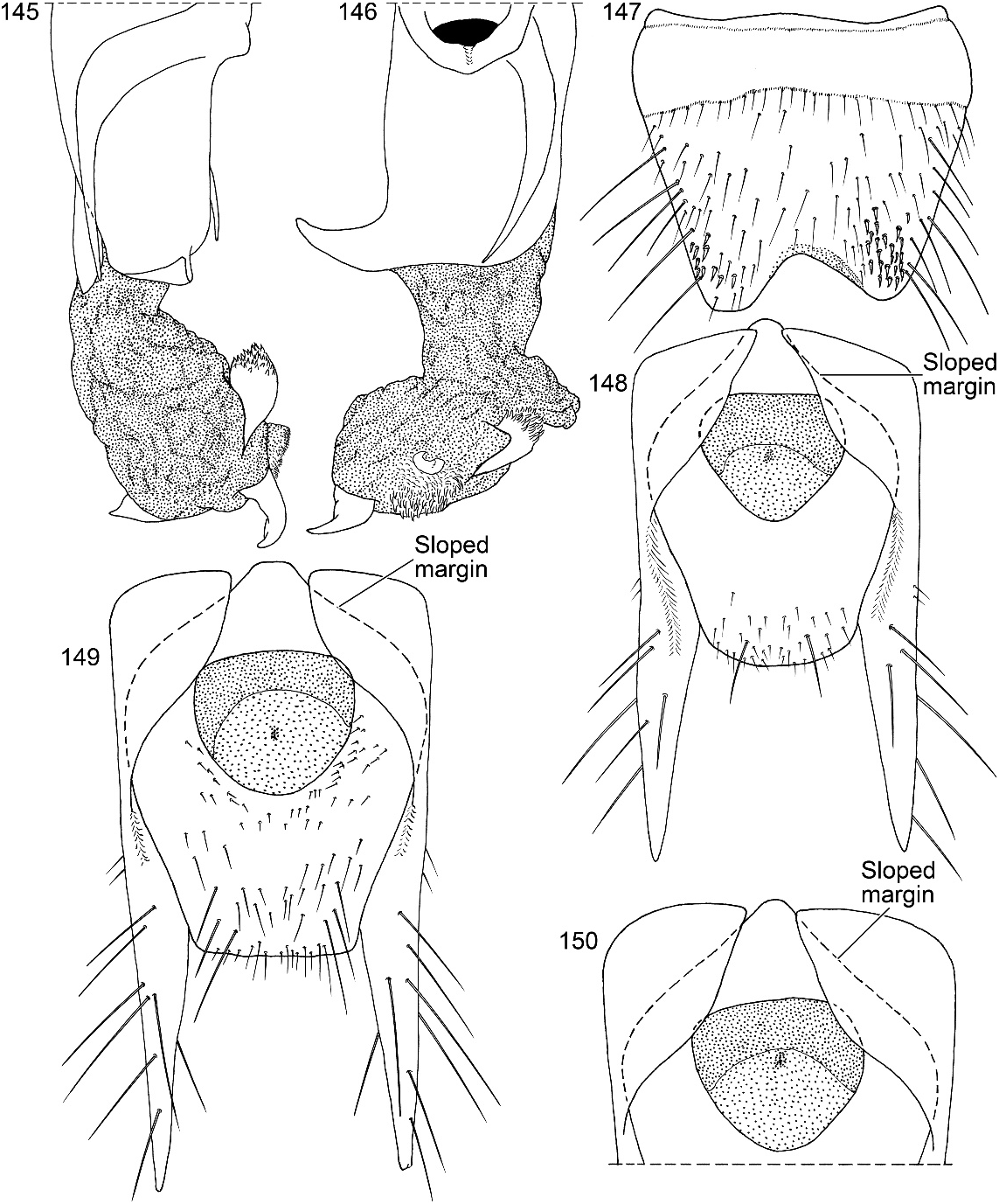
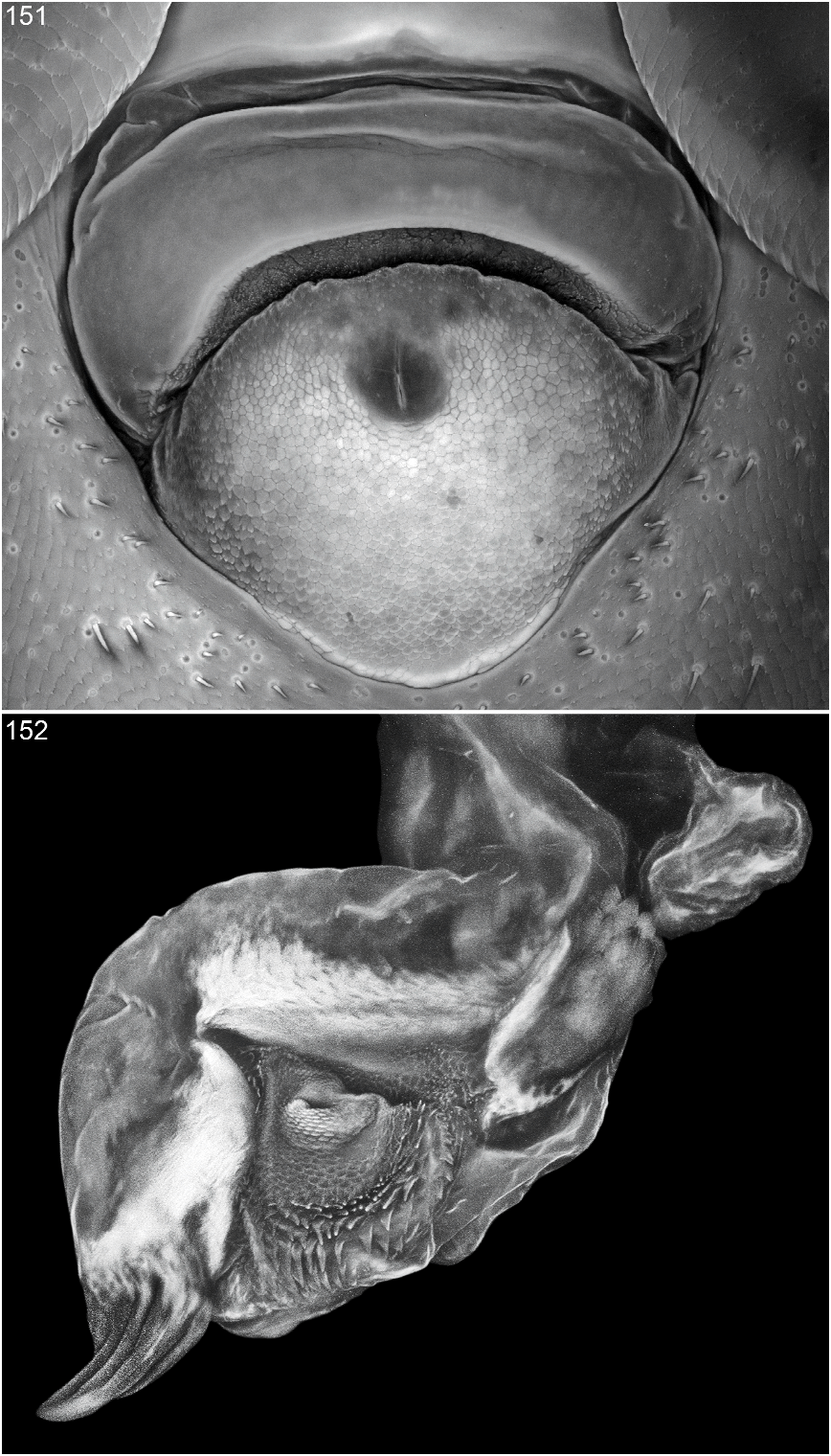
Figures 2 View Figs , 3–11 View Figs View Figs View Fig View Fig , 20–28 View Fig View Fig View Fig View Figs View Figs , 40 View Figs , 135–152 View Figs View Figs View Figs View Figs
Elytrobaeus geniculatus Sahlberg, 1847: 802 View in CoL .
Oedichirus geniculatus (Sahlberg) View in CoL
— Sharp, 1876: 338 (transferred from Elytrobaeus ). — Bernhauer and Schubert, 1912: 201 (catalog). — Blackwelder, 1944: 131 (checklist).
TYPE MATERIAL. The type series of Elytrobaeus geniculatus was not found (see Remarks below in present species account).
NEOTYPE: Designated here. ‘‘ BRAZIL, Guanabara: Corcovado Sept.–Oct. 1969 Alvarenga & Seabra/ Neotype Elytrobaeus geniculatus Sahlberg des. L. Herman, 2011/ Oedichirus geniculatus Sahlberg det. L. Herman, 2011.’’ Deposited in the American Museum of Natural History (see Remarks: Type Series below for discussion of neotype designation). (Left maxillary palpus is missing.)
TYPE LOCALITY: Brazil: Rio de Janeiro: Corcovado (22 ° 589S, 43 ° 139W).
DIAGNOSIS: Oedichirus geniculatus is one of a group of four species (also including O. clavolateralis , O. clavulus , and O. exilis ) whose males have an asymmetrical emargination of sternum VIII (figs. 94, 102, 132, 147), a stout, curved, apicoventral process extending to the right from the ventral sclerite of the aedeagus (figs. 90, 99, 130, 142), and bicolored legs (fig. 2). The surface of sternum VIII adjacent to the margin of the emargination of the four species of the group is translucent. Oedichirus geniculatus has no adornment associated with the apicolateral margin of the aedeagal ventral sclerite near the base of the apicoventral process (fig. 141), whereas near the apicoventral process O. clavulus has a peg (fig. 98), and O. clavolateralis (figs. 89, 93) and O. exilis (fig. 129) both have a laterally directed knob. The form of the apicoventral process, as depicted in the relevant figures, differs slightly among the four species (cf. figs. 90, 99, 130, 142). The labrum of the males of O. geniculatus has a tubercle at the base of the submedial denticle (fig. 136); the other species do not.
DESCRIPTION: Length: 7.7–10.0 mm. Length of head: 0.8–1.0 mm. Width of head: 1.2–1.3 mm. Pronotal length: 1.4–1.6 mm. Pronotal width: 1.1–1.3 mm. Elytral length: 1.1–1.4 mm. Elytral width: 1.2–1.4 mm.
Body concolorous pale brown (teneral specimens) to dark reddish brown to nearly black and with or without paler infusions. Legs bicolored, yellowish brown with femorotibial spot reddish brown to dark reddish brown; tibial spot smaller, paler, and less demarcated than femoral spot.
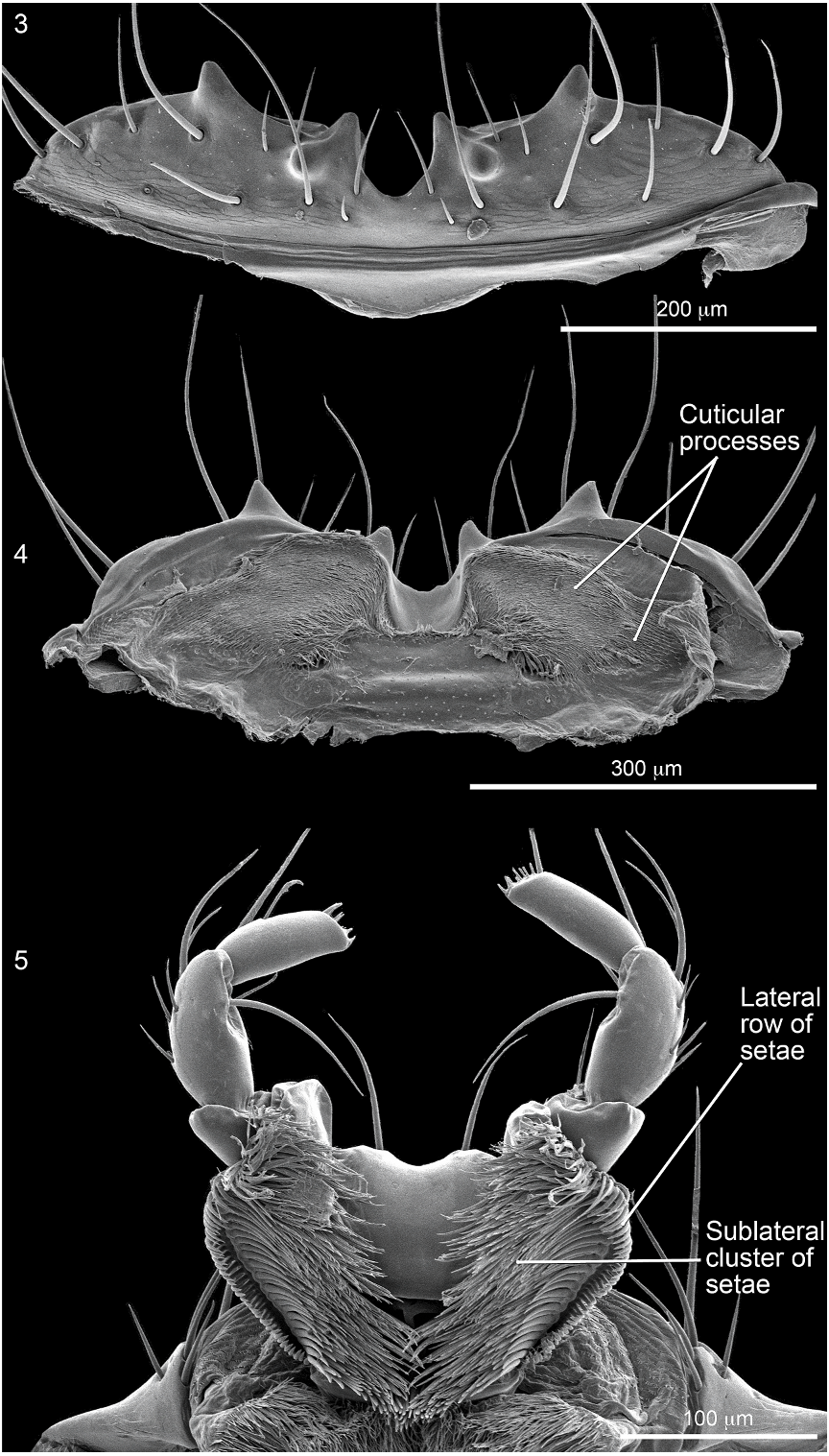
Head wider than long (HW/HL: 1.3–1.4). Frontoclypeal ridge incomplete, separated medially. Dorsal surface without V-shaped depression; surface coarsely and densely punctate; base with moderately large polished, impunctate, medial spot. Labrum quadridentate (fig. 136).
Pronotum longer than wide (PL/PW: 1.2– 1.3). Pronotum polished and with coarse, dense punctation and moderately large, impunctate spots; surface with deep, coarsely punctate, submedial groove on basal two thirds. Elytra with width equal to or slightly greater than length (EW/EL: 1.0–1.1); surface flat to feebly convex and coarsely punctate.
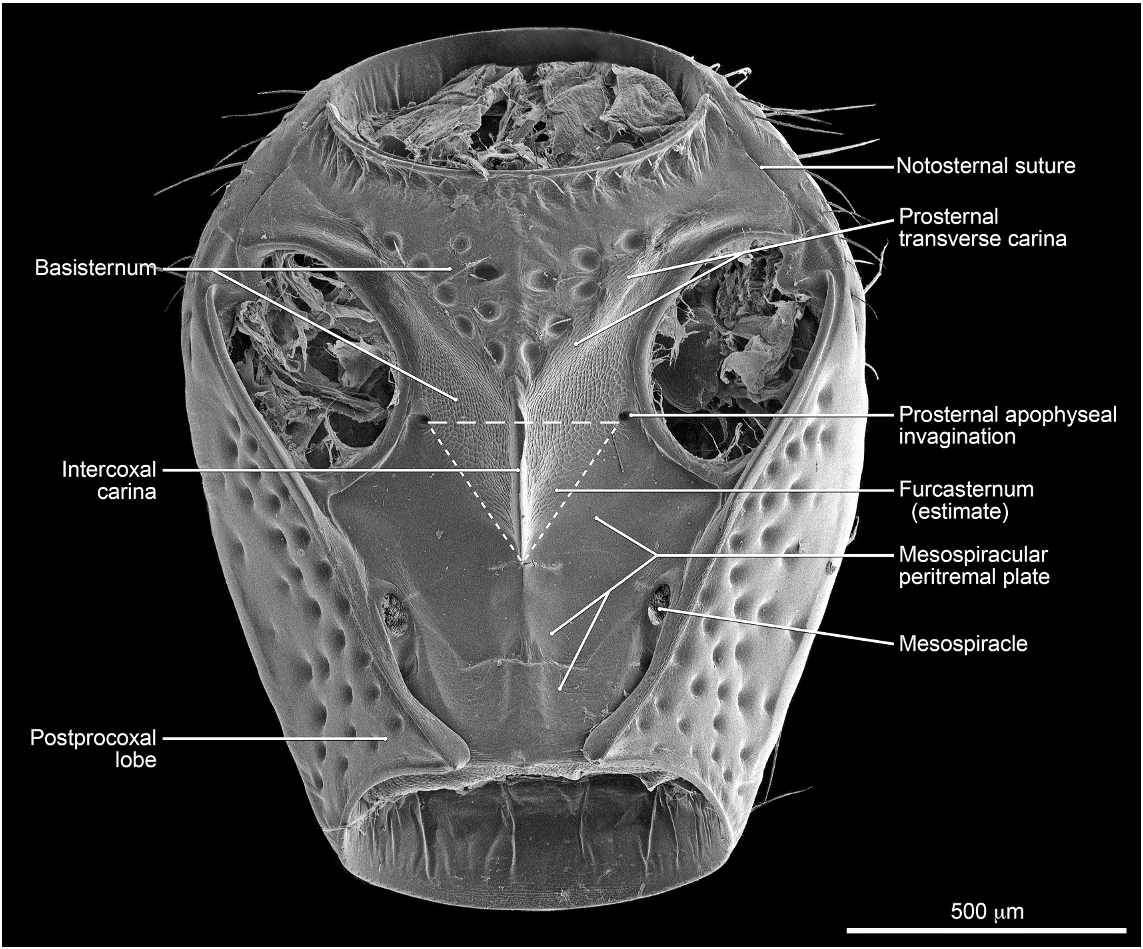
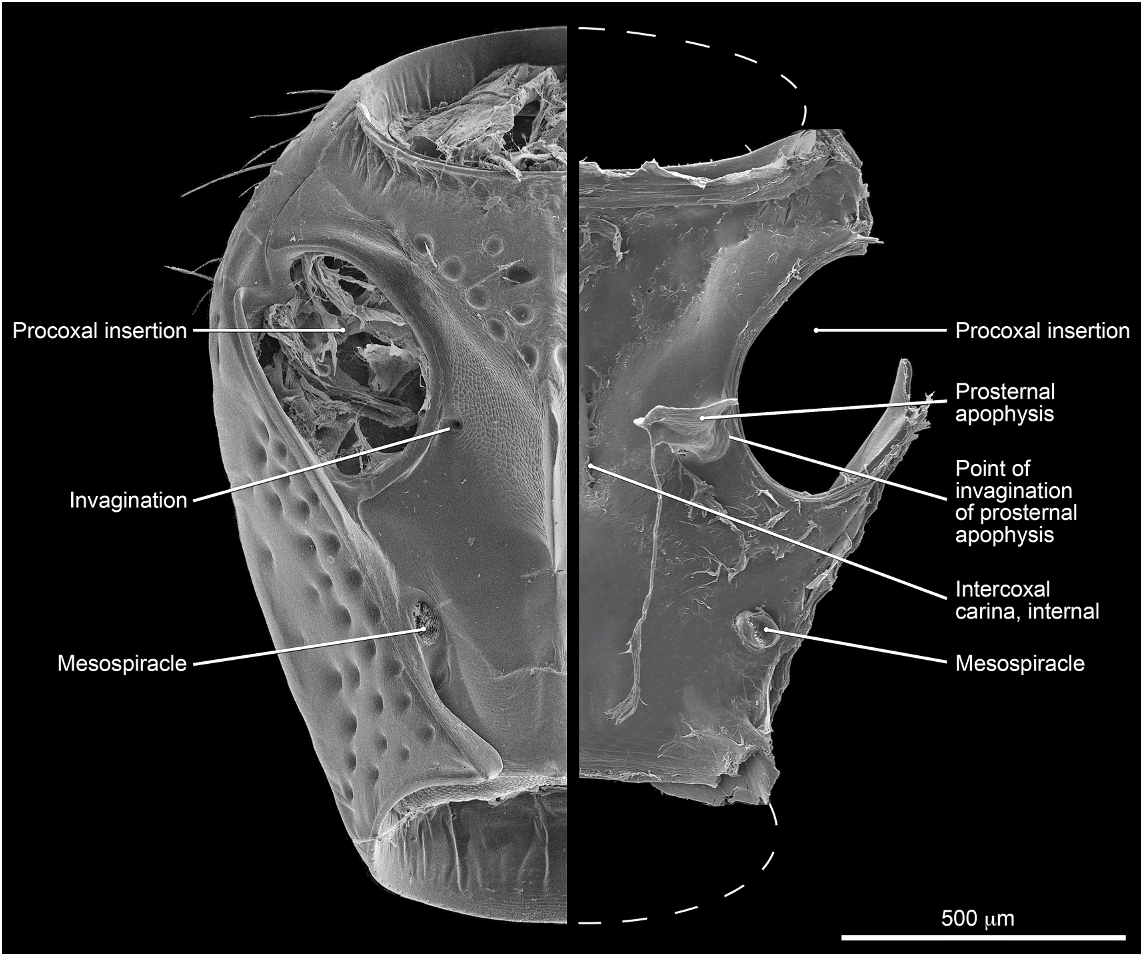
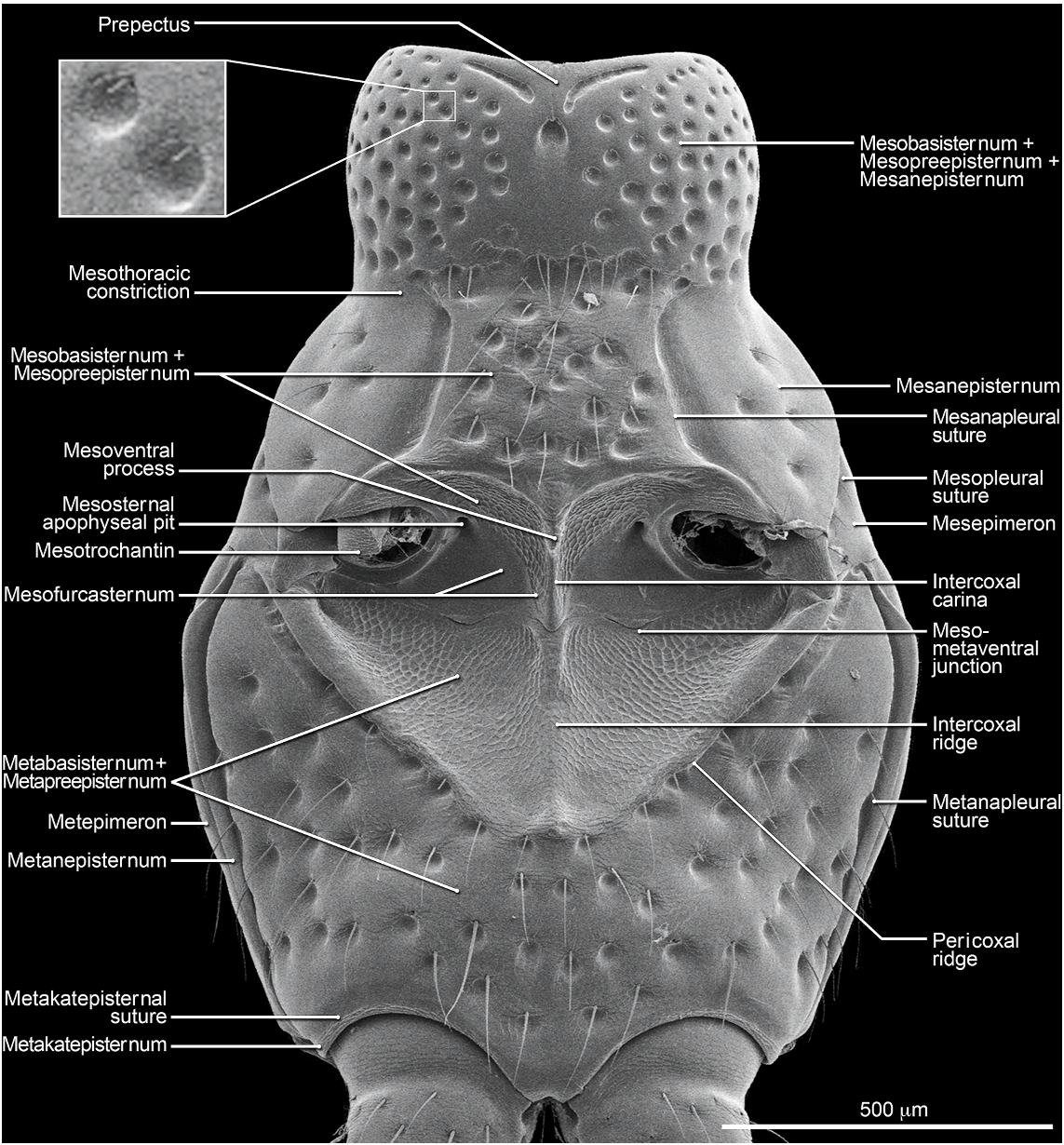
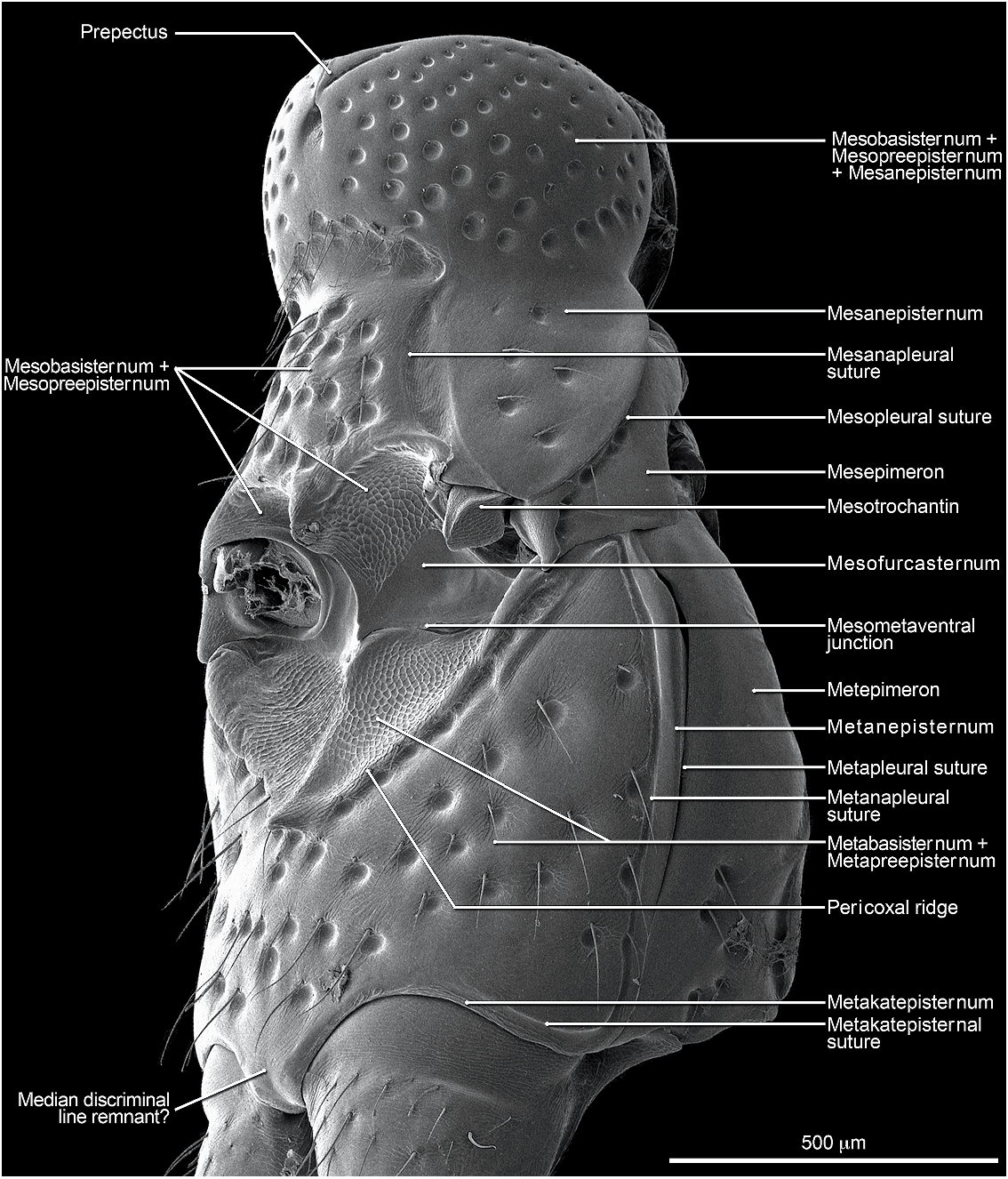
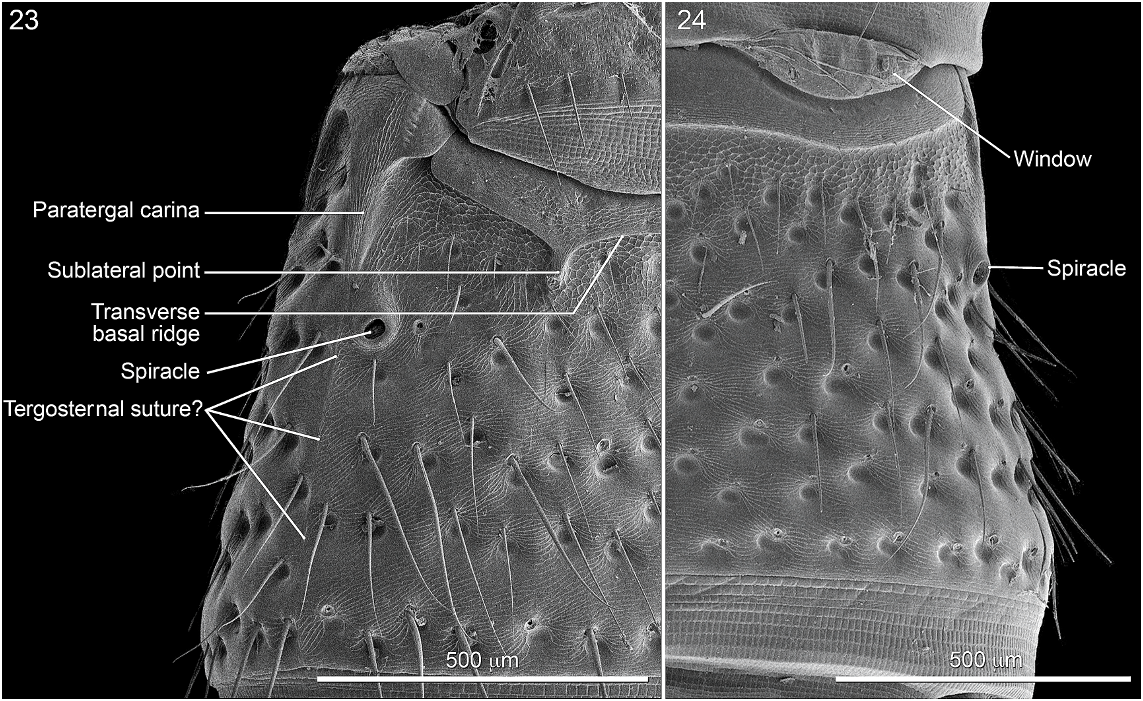
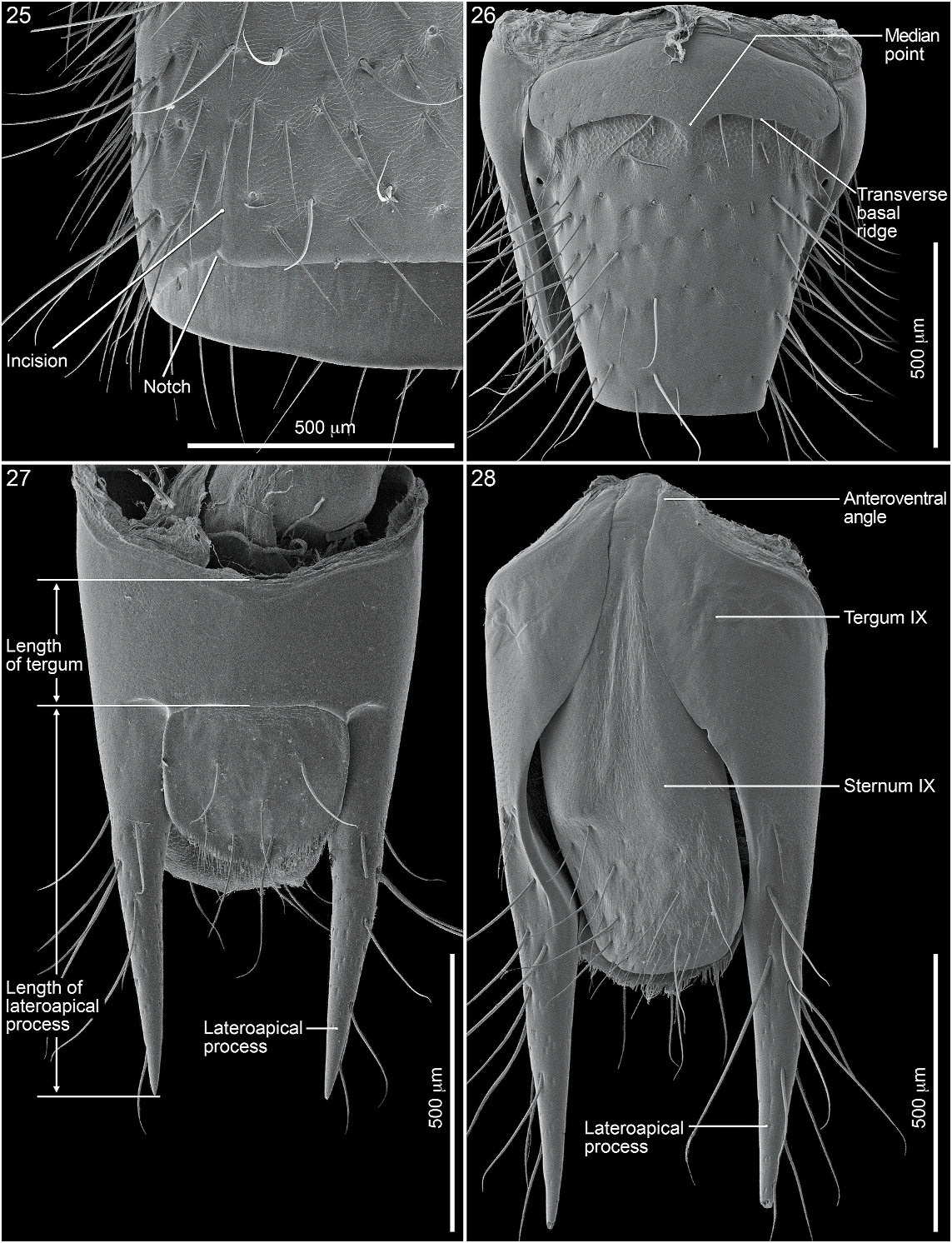
Abdomen densely and irregularly punctate; punctures not arranged in rows. Segment III without paratergite; paratergal carina present and extending to near middle of segment, but poorly developed beyond spiracle. Tergum III without median point extending from transverse basal ridge. Tergum VIII with posterior margin slightly to broadly and shallowly rounded (males) or truncate to slightly emarginate (females); transverse basal ridge feebly sinuate and with apically closed median point (fig. 26). Tergum IX (fig. 140) with lateroapical process long, more than twice as long as midbasal length (LLaP/L9 5 1.9–3.0), slightly bent ventrally, and approximately parallel to other process; ventromedial margin without posteriorly directed spur (cf. fig. 158).
MALE: Labral surface with subapical tubercle near submedial denticle (fig. 136). Sternum VI unmodified. Sternum VII with small, median, impunctate, asetate spot near posterior margin. Sternum VIII (fig. 147) with broad, deep, asymmetrical emargination of posterior margin; emargination about one sixth of length of sternum, wider than deep, with moderately broadly rounded base, and mostly to left of midlongitudinal line; right edge of emargination shorter and more steeply sloped than left; surface of margin adjacent to emargination translucent; surface with slight carina laterad of left edge of emargination; surface with cluster of short, spinelike setae laterad of each side of emargination near posterior margin and without comb or depression; transverse basal ridge unevenly sinuate and shallowly curved anteriorly and median point absent. Tergum IX with anteroventral angle extended anteriorly as robust process. Sternum IX (fig. 144) slightly asymmetrical; anterior margin narrow and strongly rounded; posterior margin wide and broadly rounded; lateral margins broadly rounded.
Aedeagus asymmetrical (figs. 141–143). Ventral sclerite with apical margin more or less truncate (figs. 142, 146), with large, flattened, curved, strongly sclerotized apicoventral process extending from right apicolateral margin (figs. 141, 142, 146); apicoventral process strongly tapered to acute apex, extending lateroventrally, and with apex bent lateroanteriorly; ventral sclerite without lobe, peg, or knob arising from right lateral edge near base of ventral process (fig. 141). Parameres moderately broad basally, flattened, tapered apically, and fused to median lobe for most of length and free of median lobe near apex; right paramere slightly longer than left (figs. 141, 143). Internal sac largely membranous (figs. 145, 146, 152); apical region near gonopore with armature on ventral surface; dorsal surface with short, stout, thornlike spine (fig. 145); ventral surface with stout, strongly curved, heavily sclerotized, grooved spine (fig. 146, 152) lateroposteriad of and at right of gonopore; gonopore surrounded by cuticular cobble and with distal patch of spiniform, cuticular processes (figs. 146, 152).
FEMALE: Labrum without subapical, submedial tubercle between denticles. Sternum VIII with broadly rounded posterior margin; transverse basal ridge irregularly sinuate and broadly curved anteriorly. Tergum IX with anteroventral angles separated (figs. 148– 150). Median gonocoxal plate anteriad of vulvar plate trapezoidal (fig. 149) to triangular (figs. 148, 150) and anterior margin nearly flat to narrowly rounded medially then gradually sloping lateroposteriorly; gonocoxal plate posteriad of vulvar plate gradually tapered to broadly rounded to truncate posterior margin. Vulvar plate embedded in anterior half of median gonocoxal plate (fig. 149). Anterior vulvar lobe (fig. 151) curved around anterior margin of posterior vulvar lobe and wider and shorter than posterior vulvar lobe; surface membranous and without adornment. Posterior vulvar lobe (fig. 151) wider than long; surface covered with cobble except near margins. Vulva oriented at slight angle to longitudinal axis.
MALE/FEMALE ASSOCIATION: The O. geniculatus complex, based on shared features of the aedeagus and sternum VIII of the male, is comprised of four species, all known only from the state of Rio de Janeiro. One of the species, O. geniculatus , is represented by 51 males from two sites in the city of Rio de Janeiro. Specimens of 32 females were collected with the 51 males and are regarded to be conspecific.
MATERIAL EXAMINED: Fifty-two males, 37 females. Brazil: ( 1 male, BMNH). Rio de Janeiro: Represa Rio Grande , August– September 1969, F.M. Oliveira (22 ° 549S, 43 ° 149W) ( 25 males, 18 females, AMNH) ; Corcovado , September–October 1969, Alvarenga & Seabra (22 ° 589S, 43 ° 139W) ( 26 males, 14 females, AMNH) ; Rio de Janeiro, Fry ( 4 females, FMNH), December 1856, H. Clark ( 1 female, FMNH) .
LOCALITIES: Corcovado is a peak that reaches 710 m elevation in a national park in the south of Rio de Janeiro. The coordinates for it and Represa Rio Grande were taken from Paynter and Traylor (1991: 162, 522).
DISTRIBUTION: This species is known only from the state of Rio de Janeiro, Brazil (fig. 40).
REMARKS: Type Series: In search of the type series of O. geniculatus , I discussed the issue with curators of collections in Helsinki, Turku, and Stockholm and examined collections and types of Oedichirus in the Natural History Museum, Field Museum of Natural History, Institut Royal des Sciences Naturelles, Museum für Naturkunde, Musée Royal de l’Afrique Centrale, and Senckenberg Deutsches Entomologisches Institut, but the type series was not found. Efforts to find it are discussed in the following paragraphs.
The R.F. Sahlberg collection is said to be in the Zoological Museum, University of Helsinki, and the Zoological Museum, University of Turku, Finland; the South American collections are specifically reported to be in Helsinki ( Horn and Kahle, 1936: 236). Type material of Elytrobaeus geniculatus was not included in a list of the types in the Helsinki collection ( Silfverberg, 1988).
In an e-mail message Jyrki Muona (Zoological Museum, University of Helsinki; 11 January 2009) wrote that ‘‘R. Sahlberg sold most of the Brazilian insects to Stockholm.’’ In response to my query Bert Viklund (Naturhistoriska Riksmuseet, Stockholm) reported that ‘‘The type material of geniculatus is not in our museum and wasn’t entered in our files and not even noted in our old Gemminger & Harold catalogue.’’ He stated that the museum has much material from Sahlberg’s South American expedition, but that the types of species he described may not have been included in the purchase. On the other hand, Muona, in a January 2009 e-mail message, wrote that all of the types of Eucnemidae described by Sahlberg were in Stockholm.
Collections at the Zoological Museum, University of Turku, also have R.F. Sahlberg specimens. Veikko Rinne ( 27 March 2009, e-mail), at the Turku collection, wrote that ‘‘This species [ E. geniculatus ] is not in our type collection where most specimens recognized as type are.’’ He continued that they have a large collection of R.F. Sahlberg’s material from Brazil among which is a box, full of Staphylinidae . Included in the box are three specimens the first of which has an old handwritten label ‘‘ Oedichirus geniculatus Sahlb. ’’ Two specimens, each with a gray rectangular label, are from Petropolis, the third, with a violet triangular label, is from Rio de Janeiro, within which the type locality of E. geniculatus is included. Based on a photograph sent to me, the specimen from Rio is most probably a female and V. Rinne wrote that the specimen is about 9 mm long ( 30 March 2009, e-mail). Sahlberg (1847: 802) cited the length as 4 lines (about 8.5 mm), so the specimen is about the right size and from the right locality. However, if this specimen were a syntype the expected generic name on the identifaction label would be Elytrobaeus , not Oedichirus . The next e-mail letter answered my concern about the identification label. The specimens in the box containing O. geniculatus had probably been lent to Bernhauer, who added the identification labels to the material (Rinne, 31 March 2009, e-mail) and thereby explaining the use of the currently accepted generic name. Elytrobaeus geniculatus was collected during Sahlberg’s first trip to Rio de Janeiro ( 12 December 1839 to 5 January 1840) and, according to his diary, the specimens he collected then were labeled with shiny green squares (Rinne, 2 April 2009, e-mail). The diary indicates that specimens with a violet triangular label, such as that on the O. geniculatus in Turku, were collected in Rio during his second trip to South America (1849–1851) (Rinne, 2 April 2009, e-mail) after O. geniculatus had been described. In summary, none of the specimens in Turku identified as O. geniculatus are syntypes and no potential syntypes were found in the collections in Helsinki or Stockholm.
Sahlberg (1847) did not indicate the number of specimens he examined for this species. The characters he provided will not separate the species from others of the genus. However, one, the unique cluster of specialized seta (5 spiniform pencil) on the apex of antennomere 11 ( Sahlberg, 1847: 803 ‘‘ultimo oblongoovato, apices spina acuta armato…’’), does permit generic assignment (see fig. 9). The locality from which the species was collected in Rio de Janeiro was clearly identified: ‘‘cum omnia in vicinitate urbis Rio Janeiro, atque qua maximam partem in tractu montuoso (Corcovado) capta sunt’’ ( Sahlberg, 1844: 501). Although we know the size, color, generic assignment, and original collecting site for the type series, and although no specimens of the type series were found, is selection of a neotype justified?
To designate a specimen as a valid neotype, the Code ( ICZN, 1999, article 75.3) explicitly qualifies that evidence be presented that the neotype be ‘‘consistent with what is known of the former namebearing type from the original description…’’ (Article 7.3.5) ‘‘and that the neotype [come] as nearly as practicable from the original type locality…’’ (Article 7.3.5). Beyond the character of antennomere 11, only the originally stated size and locality help narrow the choice of a neotype from among the four species known in the state of Rio de Janeiro. In the present work only one species was collected at Corcovado and the range of the length of that species also overlaps the stated size of O. geniculatus . Although the remaining three species might be found at Corcovado, none were. Two were found well outside of the city limits of Rio de Janeiro in Petropolis ( O. clavulus ) and Silva Jardim ( O. exilis ) and the third was from an unspecified part of Rio de Janeiro ( O. clavolateralis ). Each of the remaining three species is within range of the size of O. geniculatus , but only one sample encompassed the size of the original specimen. I regard the specimens studied herein from Corcovado to be O. geniculatus as they provide the best fit for the few known facts.
The conditions required for designation of a neotype are that (1) no extant name-bearing specimen is believed to exist, (2) a type is needed to objectively define the nominal taxon ( ICZN, 1999: Art. 75.1), and (3) there is an expressly stated exceptional need to designate a neotype ( ICZN, 1999: Art. 75.3).
When designating a neotype authors generally state that the original type series is lost and that a neotype is needed to objectively fix the identity of the name. In addition to noting that the primary type is lost, two other reasons cited for neotype designation are to fix the identity of nomina dubia ( Assing, 2008: 1239) and to fix a name with a long history of being identified in a manner that is contrary to the original description ( Frisch and Herman, 2008). Another reason would be to fix the name of a species in a cluster of subtly differentiated species such as is the case for O. geniculatus , which, without examination of the aedeagus, is easily confused with O. clavolateralis , O. clavulus , and O. exilis . The type of O. geniculatus is lost; a name-bearing type will provide an objective basis for identification.
The conditions set forth in Article 75.3 are herewith addressed. A neotype is needed for O. geniculatus to provide an objective basis for its separation from three similar species; the diagnostic features are presented in the preceding diagnosis and description for the species; the label data are presented above in the paragraph ‘‘ Neotype,’’ and my neotype label is attached to the specimen pin; there is ample evidence presented in preceding paragraphs that the original type series cannot be found and is therefore presumed lost; the specimen selected is from the same locality and is approximately the same size; the specimen is deposited in the American Museum of Natural History. A neotype is hereby and herein designated for Elytrobaeus geniculatus Sahlberg.
R.F. Sahlberg’s Types: The question of the depository of the syntype (s) of E. geniculatus concerns more than the location of the type material for one name. Sahlberg described 19 species of Staphylinidae in his 1844 and 1847 articles on beetles from Rio de Janeiro. According to Horn and Kahle (1936: 236) that material should be in the Zoological Museum of the University of Helsinki. In the list of staphylinid types for that collection ( Silfverberg, 1988) only seven are reported; 12 were not mentioned. Among the missing are (arranged by subfamily): Falagria brasiliensis , Oxypoda lapidiola , Bolitobius gilvipes , Tachinus tropicus , Tachyporus albicornis , Tachyporus fulvus , Belonuchus vestitus , Quedius nigritulus , Dibelonetes biplagiatus , Elytrobaeus geniculatus , Taenodema villosa , and Oxytelus brasiliensis . Of the foregoing, Dibelonetes biplagiatus is the type species for the genus.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oedichirus geniculatus (Sahlberg)
| Herman, Lee H. 2013 |
Elytrobaeus geniculatus
| Sahlberg, R. F. 1847: 802 |