
Oedichirus Erichson
|
publication ID |
https://doi.org/10.1206/816.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF8794-7D12-D146-FF59-5507FB090369 |
|
treatment provided by |
Felipe |
|
scientific name |
Oedichirus Erichson |
| status |
|
Oedichirus Erichson View in CoL
Oedichirus Erichson, 1839: 29 View in CoL (published without included species; for first included species, see Erichson, 1840: 685). Type species: Oedichirus paederinus Erichson View in CoL , fixed by Erichson (1840: 685) by subsequent monotypy. (The following subsequent references for Oedichirus View in CoL include only those pertaining to the New World fauna. For a more extensive list, see Herman, 2010: 36 View Cited Treatment .) — Erichson, 1840: 684 (first included species: paederinus View in CoL ; characters). — Sharp, 1876: 338 (first two New World species described in Oedichirus View in CoL ). — Blackwelder, 1944: 131 (checklist of species of Latin America). — Blackwelder, 1952: 269 ( type species). — Navarrete-Heredia et al., 2002: 293 (characters in key to genera for Mexico; general notes; unnamed species in Mexico). — Herman, 2010: 36 View Cited Treatment (characters; list of species included; phylogenetic position).
Elytrobaeus Sahlberg, 1847: 801 View in CoL . Type species: Elytrobaeus geniculatus Sahlberg View in CoL , fixed by monotypy.
— Gemminger and Harold, 1868: 629 (catalog). — Fauvel, 1875a: xvii (5 1875b: 219) (synonym of Oedichirus View in CoL ). — Sharp, 1876: 338 (synonym of Oedichirus View in CoL ). — Bernhauer and Schubert, 1912: 201 (synonym of Oedichirus View in CoL ). — Cameron, 1931: 25 (synonym of Oedichirus View in CoL ). — Blackwelder, 1952: 147 ( type species).
Oedichiranus Reitter, 1906: 263, 264 (subgenus). Type species: Oedichirus ( Oedichiranus) dimidiatus Reitter , fixed by monotypy.
— Herman, 2010: 38 (synonym of Oedichirus ).
DIAGNOSIS: Oedichirus can be separated from all other genera of the Procirrina by the apical, spiniform pencil of antennomere 11 (fig. 9) and the basal fusion of tergum and sternum VII (fig. 25). Only Oedichirus and Palaminus have windows near the anterior margins of terga and sterna III to VII (fig. 24) and a dense cluster of setae on the ventral surface of protarsomere 5 (fig. 7). Tergum and sternum VII are fused in Palaminus , but fused in Oedichirus from the base to near the apex then separated by a short incision (fig. 25). The surface of the abdomen of Oedichirus is punctate (figs. 23, 24) and lacks the imbricate macrosculpturing of Palaminus (fig. 31). The long, thick, lateroapical elytral seta found in Palaminus (fig. 30) is absent in Oedichirus (fig. 8). The posterior edge of the elytra lack a row of setae (figs. 1, 2, 8) and the basal four protarsomeres are inflated in Oedichirus (figs. 6, 7).
DESCRIPTION: Head broad; postocular lateral margin short and straight (figs. 1, 2); basal margin of head extending anterolaterally from neck and with ridge extending laterally from neck to near eye; ridge acutely angulate laterally. Eye length greater than postocular length of head. Dorsal surface of head with umbilicate punctation ( Herman, 2010: fig. 15); frontoclypeus polished, impunctate, and with median impression. Antennomere 10 and 11 of subequal length; antennomere 11 with spiniform process apically (fig. 9). Labrum (fig. 3) with moderately deep median emargination of anterior margin; anterior margin with one, two (fig. 3), or three pairs of denticles. Mandible (figs. 2, 137) with one denticle near middle. Maxillary palpus (fig. 138) with fourth segment shorter to slightly longer than second and enlarged, robust, moderately compressed, and covered with fine, dense pubesence.
Pronotum longer than wide (figs. 1, 2, 10); widest near anterior third and with lateral margins strongly convergent anteriorly and gradually convergent posteriorly; punctation umbilicate, sparse to dense, absent from midline medially and posteriorly, present or absent from midline anteriorly, and confused, arranged in curved rows, or grouped in clusters. Pronotal marginal ridge present, complete, and well developed. Mesospiracular peritremal plate (fig. 10) large, strongly sclerotized, fused anteriorly to furcasternum and laterally to hypomeron. Procoxal cavity closed posteriorly by mesospiracular peritremal plate (fig. 10). Elytra (figs. 1, 2) shorter than pronotum; surface with uniform punctation; posterior margin without row of setae on edge; apicolateral corner without long prominent seta (fig. 8).
Protarsomeres (figs. 6, 7) 1 to 4 inflated and with crescent-shaped slit on ventral surface; slit bordered by setae; ventral surface without setose pad; tarsomere 4 not expand- ed beneath 5 and apical margin entire, not emarginate; tarsomere 5 densely setose ventrally (fig. 7). Mesotarsomeres and metatarsomeres ( Herman, 2010: fig. 50) 1 to 4 slender and each with long first tarsomere; tarsomeres 1–3 pubescent ventrally, but without setose pad; tarsomere 4 extending beneath 5, apical margin entire to slightly emarginate, dorsal surface deeply impressed medially, and ventral surface with dense cluster of long setae.
Abdominal segments punctate and without imbricate macrosculpturing (cf. figs. 23 and 31). Abdomen with oval, membranous ‘‘window’’ (fig. 24) in intersegmental membrane at anterior margin of terga and sterna III–VII. Segment III with paratergite or paratergal carina (fig. 23); tergum and sternum separated by tergosternal suture and paratergite in some species or by paratergal carina and feeble groove or line (fig. 23) in others. Segments IV to VII without paratergites or paratergal carinae and with tergum and sternum on each segment fused. Segment VII with tergum and sternum fused basally, posterior margin with notch on midlateral side and with short incision extending anteriorly from notch (fig. 25). Sternum III with midlongitudinal carina; transverse basal ridge with sublateral carina extending posteriorly ( Herman, 2010: fig. 22). Tergum III with two or three wedge-shaped points extending posteriorly from transverse basal ridge; sublateral point present (fig. 23), medi- an point present (as shown for tergum or sternum VIII, figs. 26, 73) or absent. Tergum VIII with (figs. 26, 166) or without median point extending from transverse basal ridge. Tergum IX fused middorsally (fig. 27); posterior margin truncate and deeply emarginate medially; lateroapical process (fig. 27) long, slender, slightly (fig. 158) to strongly curved (fig. 168) ventrally, and extending beyond apex of tergum X. Sternum IX of male slightly (figs. 51, 86, 109) to strongly (fig. 117) asymmetrical; base wide (fig. 117) to narrow (fig. 144). Tergum X (fig. 140) present and nearly entirely exposed; apical margin curved to slightly produced medially.
Male with sterna VI and VII modified or unmodified; sternum VIII modified. Aedeagus with parameres present (figs. 103, 104, 161, 162); basal piece absent.
Female with sterna VI–VIII unmodified. Median gonocoxal plate undivided (fig. 29. Vulvar plate with two lobes (figs. 29, 111, 157, 196). Vulva embedded in posterior vulvar lobe and surrounded by membrane (figs. 29, 64, 196).
DISTRIBUTION AND HABITAT: Most species of Oedichirus are found in tropical and subtropical regions of the world; a few are
known from temperate regions. The New World species are known from Mexico to southern Brazil, northern Argentina, and Bolivia, but have been captured in few countries and at few sites (figs. 39, 40). Most of the previously described species are found in Brazil and one is from Costa Rica, but herein species are reported from the Dominican Republic, Mexico, Ecuador, Peru, Bolivia, Argentina, and perhaps Nicaragua.
Little has been published and few data are on specimen labels that indicate the habitat of the New World species, but labels record specimens to have been collected from shrubs, trees, and leaf litter.
In the Old World, Oedichirus is widespread and represented by many species. Subsaharan Africa and Madagascar are densely populat- ed by the genus. Four North African species occur along the southern edge of Europe in Spain, Italy, and Crete and two others have been reported from countries adjacent to the Mediterranean and from there the genus is found from India to Japan and through Indonesia to Australia. One species reaches Central Asia ( Tajikistan and Afghanistan), one species is reported from Tasmania, and two are from New Caledonia. Old World species have been collected from leaf litter on the forest floor, moist ground litter, and in swamps, near streams and springs, and from vegetation by beating shrubs and branches of trees, and epiphytes and are found in lowland forests and in montane forests to as high as 3500 m. At least 10 African species have been collected at or above 3000 m elevation (see Fagel, 1971: 194, 237, 241, 243–245, 252, 266, 287, 291).
MORPHOLOGY OF OEDICHIRUS
The following summary of the morphological variation of Oedichirus is based primarily on New World species, includes some speculation, and introduces a few new structures and terms. So the reader can better understand the rationale behind the terms used herein for the lateral and ventral pterothoracic sclerites of Oedichirus , a discussion of two hypotheses concerning the evolution of the pterothorax is provided.
COLOR: Most species of the genus are varying shades of reddish brown to reddish black to black, different segments of the body may be paler or darker and the legs of some species are bicolored. Like species in some other staphylinid genera, some species of Oedichirus mimic a color pattern common to many species of Paederus , one in which the head, elytra, and apical abdominal segments are black and the remaining regions are orange to reddish orange. Among the Old World Oedichirus at least 30 species have this color pattern, but there may be others among those I have not examined. All but four of the 30 species are African; the others are from Japan (2), India (1), and Central Asia (1). One of the Paederus -like African species, O. paederinus Erichson , is also from Europe and is the type species of the genus (fig. 1). All of these mimetic species, except O. paederinus , have fully developed elytra and probably wings and may be capable of flight; all of them lack the pronotal marginal ridge. Coiffait (1978: 330, 332) reported the same color pattern in O. simoni , which I’ve not seen, from the Mediterranean region. I have seen no Australian species so colored, but although MacLeay (1873: 147) reported the head and apical three abdominal segments for O. paederoides as black and the thorax and basal abdominal segments reddish, he did not describe the elytra. In the New World I have seen no species that as closely resemble the color pattern of Paederus as do the mimetic Old World species. Those with the closest resemblance are O. bullaglaber in which the head and prothorax are black and the remainder of the body is reddish brown and O. neotropicus in which the body is reddish brown and the apical abdominal segments are black. The legs of 12 New World species are bicolored.
HEAD: Excluding the eyes, the head is more or less quadrate in dorsal view in many species, the dorsal surface is densely to sparsely punctate, and the punctures are large and deep to relatively small and shallow (figs. 1, 2). Some species have almost no dorsal punctures and all have impunctate spots. The ventral punctation is sparse and shallow. Trichobothria are absent. The frontoclypeal region, usually polished and depressed medially, has a low frontoclypeal ridge extending medially from near the antennal insertion and is complete or incomplete. The basal margin of the head is broadly and gradually rounded from the eye to the neck (fig. 1) or has a carina extending from the neck laterally to near the base of the eye where the carina is strongly angulate (fig. 2). The neck is about half to two thirds as wide as the postocular width of the head. Separating the head and neck is a nuchal constriction, the dorsal portion of which is termed the nuchal groove, followed posteriorly by the nuchal ridge on the neck. The nuchal groove lacks longitudinal carinae. On the ventral surface of the neck the postoccipital suture is evident as a well-developed ridge, that extends anteriorly as the gular sutures, which are moderately to poorly developed or absent (see Herman, 2010: fig. 16). The gular sutures are separated, shallow grooves that when feebly developed are evident only as slight impressions and when absent leave no impressions on the surface, but are visible through the cuticle as dark lines. The eyes are more or less protuberant from the sides of the head and lack setae, each facet is nearly flat and only feebly raised, and the posterior margin of the eye is not emarginate.
Most antennomeres are not notably modified, but 11 has a long, spiniform apical pencil (fig. 9) that, with a light microscope, appears to be a spine or tightly packed cluster of setae. Unique to Oedichirus , the spiniform pencil was noted by Sahlberg (1847: 303), then by Sharp (1876: 338, 339), who was unsure of what it was. SEM images show a group of three wide, flattened processes that are apparently modified setae. Each seta is wider than thick, tapers apically, has longitudinal ridges extending for most of the length, and three apical cuticular processes. This spiniform pencil is partially withdrawn into the apex of the antennomere in some individuals because the apex is more or less membranous and collapses in dried specimens. The function is unknown, but the pencil may serve to wick chemicals produced by antennal glands to the surface for dispersal. Secretory glands have yet to be discovered.
While the anterior margin of the labrum of most New World species has four denticles, O. optatus has six, and O. echinatus , O. lunatus , and O. procerus have two. One species, O. geniculatus , has four denticles and a small, dentiform, subapical tubercle near the base of the submedial denticle in males (figs. 3, 136) and O. echinatus has a minute tubercle at the edge of the anterior margin adjacent to the median emargination. The epipharynx lacks setae, has a deep median groove, and the median region has a dense, transverse mat of cuticular processes (fig. 4).
Typical of most staphylinids the labium has three, undistinguished palpomeres that are increasingly slender from the thick, basal article (fig. 135). The anterior margin of the hypopharynx has a pair of rounded to truncate lobes. The hypopharynx has, on the anterior margin, a pair of rounded to truncate lobes, but lacks clusters of spinelike setae; the lateromedial surface is densely covered with cuticular processes and bordered laterally by a row of setae (fig. 5).
The maxilla has four palpomeres (fig. 138). The basal palpomere is small and the second and third long, slender, and slightly expanded apically. The fourth palpomere is greatly enlarged and the sides are covered with fine pubescence while the apical surface is covered with sensory structures similar to sensilla basiconica (see Herman, 2010: fig. 30). This enlarged, pubescent palpomere is characteristic of species in the Procirrina and Pinophilina and SEM images may reveal significant variation of potential diagnostic or phylogenetic interest, but at present insufficient material exists for most species or genera to determine this.
The mandibles are long, slender, have only one simple, apically acute denticle near the middle (fig. 137), and the prostheca is a simple, basal cluster of cuticular processes.
PROTHORAX: The prothorax (figs. 1, 2) is tapered posteriorly from about the anterior third. The dorsal surface of the pronotum is moderately to strongly convex and coarsely punctate in most species. The punctures are unevenly distributed in most species, but among the New World ones, O. dilophus has smaller, shallower, marginally more uniformly arranged punctures than do the others. All of the species have scattered, polished, impunctate spots. Trichobothria are absent. The notosternal suture is present and short and separated from the marginal ridge (fig. 10). The basisternum has coarse, deep punctures, each bordered by a seta near its anterior margin (fig. 10; the seta of many punctures missing or broken). The postprocoxal lobe (fig. 10), the posteroventral part of the hypomeron, is long, tapered, and covered with dense, deep, coarse, asetate punctation.
The pronotal marginal ridge is present and complete in New World species. However, among Old World species the ridge is present and complete or divided or absent; based on the three states Fagel (1971: 129–130) arranged the African species into three groups. The marginal ridge is present in most staphylinids and most paederines and, as pointed out by Lecoq (1986: 9), its presence must be considered ancestral. Whether there is phylogenetic significance in variation of the ridge is yet to be tested. The ridge of most, perhaps all, species of Oedichirus is margined by a row of punctures along the dorsal edge.
Except for the procoxal cavities and spiracular openings, the entire ventral surface of the prothorax is covered by a large, undivided sclerite in Oedichirus . This compound, prosternal/mesotergal sclerite, which incorporates four strongly sclerotized plates, the basisternum, furcasternum, and two mesospiracular peritremes, was discussed in more detail previously ( Herman, 2010: 16–17). The basisternum extends between the notosternal sutures and from the anteroventral prothoracic margin posteriorly to the two prosternal apophyseal pits, which mark the anterior edge of the furcasternum (5 sternellum). The large sclerite behind the furcasternum, the mesospiracular peritremal plate, is formed by the enlarged, fused mesospiracular peritremes and covers the surface between the hypomera and the area from the procoxal cavities and furcasternal region to the posterior margin of the ventral surface (figs. 10, 139). Since spiracles are part of the dorsum ( Snodgrass, 1935: 71– 72, 249 and figs. 35, 137) and the tergum is sclerotized dorsum, the spiracular peritremal plate is part of the mesotergum. Between the procoxae is a large, midlongitudinal intercoxal carina, extending posteriorly from the prosternal transverse carina and ending at the midposterior edge of the mesospiracular peritremal plate.
PTEROTHORAX: The elytra of most species of Oedichirus are densely and coarsely punctate. Each puncture has a seta near the anterior margin. The dorsal surface, the disc, of the elytra of most species of Oedichirus is slightly convex to flat. For three species, O. boehmi , O. ohausi , and O. procerus , most of the disc is moderately to strongly concave and the beetle has a swayback appearance. The elytral disc of three other species, O. brunneus , O. hamatus , and O. neotropicus , is weakly concave and the elytra of O. optatus are flat to slightly concave. The posterior margin of the conjoined elytra is emarginate (figs. 2, 8) and lacks a row of setae on the edge (fig. 8). The elytra lack an epipleural ridge. Most species of Oedichirus , and all of the New World ones, are flightless with the concomitant reduction of the mesothorax and metathorax as well as further shortening of the elytra, reduction of the epipleuron, and loss of the humeral angles. The elytra of the flightless species are shorter than the prothorax. The flying species of Oedichirus have fully developed elytra that are longer than the prothorax and are scattered in Africa and Madagascar and across Asia to Indonesia and Japan.
Hypotheses of origin of lateral and ventral sclerites of pterothorax: The pterothoracic ventral and lateral sclerites of adult staphy- linids were referred to as epimeron, episternum, and sternum for both segments by Blackwelder (1936) and most taxonomists have similarly referred to these sclerites. Naomi (1988) used epimeron, anepisternum, and basisternum + preepisternum, respectively, for the same sclerites. Thayer (2005) return- ed to the use of epimeron and episternum, but dispensed with sternum and basisternum + preepisternum and used instead ventrite for the large, median ventral sclerite. Lawrence et al. (2011: 33) briefly discussed the replacement of the term ‘‘sternum’’ and adopted the view that the external sterna of the pterothorax have been lost. Because of the revised interpretation of the pterothorax, the following paragraphs briefly summarize the infrequently discussed rationale for the changes of the pterothoracic vocabulary.
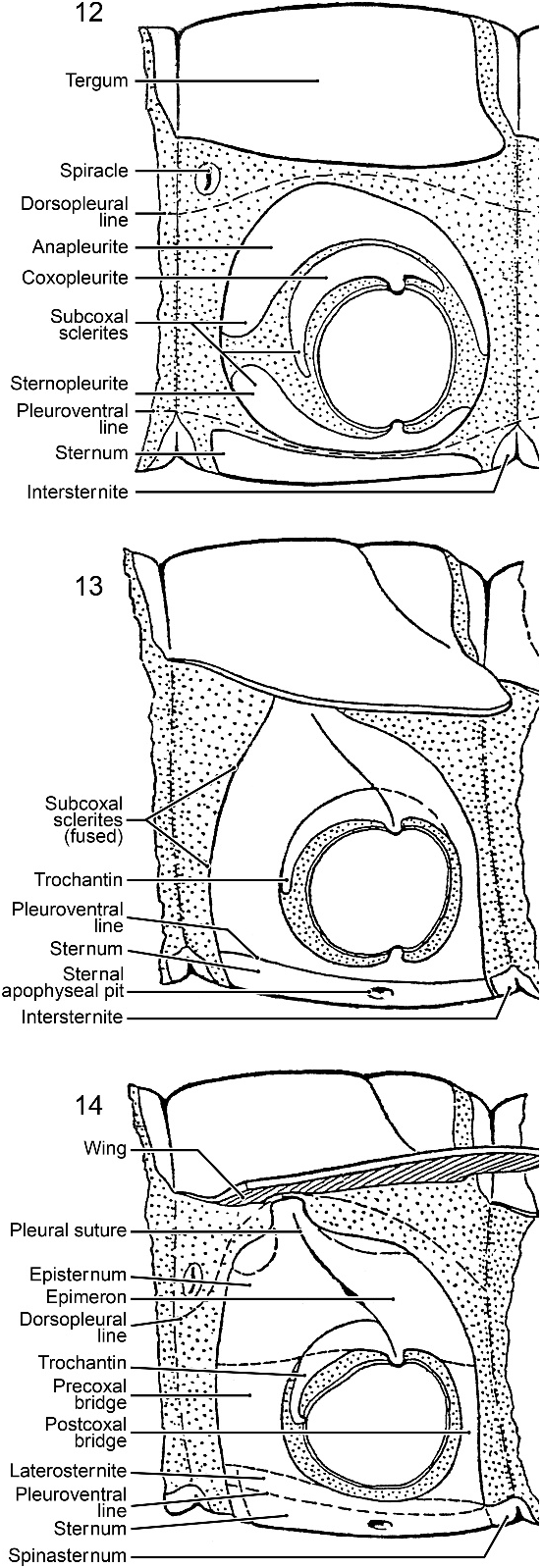
Snodgrass (1935: 161–166, fig. 88) considered the pleural sclerites to be derived from subcoxal sclerites ( figs. 12–14 View Figs ) of limb bases and that three main sclerites comprised the eventual sclerotized pterygote pleuron, two above the coxa, the anapleurite and coxopleurite, and one below, the sternopleurite ( fig. 12 View Figs ). In pterygotes, to strengthen the thoracic walls for flight ( figs. 13–14 View Figs ), the anapleurite fused with part of the coxopleurite, enlarged to cover the dorsal, anterior and posterior pleural regions, and was divided by an invagination, the pleural suture, to form an internal apodeme, the pleural apophysis, thereby creating the episternum anteriorly and the epimeron posteriorly. The remainder of the coxopleurite became the trochantin. He also named two sclerotized ‘‘bridges’’ ( figs. 14 View Figs , 18), the precoxal bridge (5 precoxale) and
postcoxal bridge (5 postcoxale), in front of and behind the coxa, that extend toward the sternal region from the episternum and epimeron, but he elaborated no further on them. The third subcoxal sclerite, the sternopleurite (5 laterosternite) ( figs. 14 View Figs , 18), fused to the sternum (and the coxal bridges, according to his figure 91B (5 fig. 14 View Figs herein), although Snodgrass did not state that). The laterosternite was little mentioned by Snodgrass or subsequent investigators. Finally, the episternum and epimeron each divided resulting in the dorsal anepisternum and anepimeron and the ventral katepisternum and katepimeron; the preepisternum is an anterior division of the anepisternum ( Snodgrass, 1935: 184). Matsuda (1963: 62–67) supported Snodgrass’s ideas, but suggested slight modifications concerning the articulating condyles.
Moving from the pleural to the ventral side, Snodgrass (1935: 77–78, 166–172) regarded the thoracic sternum or eusternum to be a composite sclerite that developed from sternal and pleural elements (figs. 15–18). Included among the sclerites are the primitive sternum medially, the intersegmental sclerite (spinasternum) posteriorly, and the sternopleurite laterally (5 his laterosternite) (fig. 18). The primary sternal sclerite has a pair of invaginations, the sternal apophyseal pits, the origin of the sternal apophyses (figs. 16–18). The regions anterior and posterior to the sternal apophyses are the basisternum and furcasternum (5 sternellum) (fig. 17), and along the anterior margin the presternum separated from the basisternum (fig. 18). Despite the composite nature of the ventral sclerite, Snodgrass (1935: 78) referred it as ‘‘sternum’’ and that designation has been followed by innumerable entomologists.
Weber (1928), studying species of Papilio , Zygaena , Sphinx , and Sialis , proposed that the external sterna of the pterothorax were
+
lost in holometabolous insects by invagination along a midlongitudinal line during postembryonic development and that the pterothoracic venter was comprised of pleural elements. Snodgrass (1935: 170) remarked only that Weber’s hypothesis that the sternal apophyses were between the lateral edge of the primary sternum and the medial edge of the subcoxal laterosternite was not always true.
Ferris and his students ( Ferris and Rees, 1939; Rees and Ferris, 1939; Ferris, 1940a), although not citing Weber, wrote that external pterothoracic sterna do not exist in Panorpa nuptialis Gerstaecker (Panorpidae) , Tipula reesi Alexander (Tipulidae) , and Plega signata (Hagen) (Mantispidae) . Campau (1940), a student of Ferris, thought the metathorax of Chauliognathus pennsylvanicus (De Geer) (Cantharidae) lacked a sternum, but that mesosternal elements were present (fig. 19). The pterosternal apophyses of the four species are close together (except for those of the mesothorax of C. pennsylvanicus ) and the ventral sclerite has a fold or furrow (discrimen) or median line (discriminal line [present only on the metaventral sclerite of C. pennsylvanicus ]) that they interpreted as indicating the point of invagination of the entire basisternum and the meeting point of the subcoxal sclerites (fig. 19). The area on each side of the discrimen or discriminal line was referred to as the discriminal area (5 preepisternum, fig. 19) and regarded to be of subcoxal origin ( Ferris, 1940a: 36–37). Ferris (1940b: 88) extended this view to state that ‘‘in the vast majority of insects—probably 95 percent of them, … the sternal apophyseal pits of the pterothoracic segments are set close together on the midline of the body.’’ He considered close proximity to be the ‘‘primitive position’’ and that any more lateral position of the apophyses was correlated with the loss of ‘‘the ventral subcoxal arc’’ (5 sternopleurite?; preepisternum?), in which case the resulting sclerite between the apophyses was truly sternal. In another article, Ferris (1940a: 36) stated that in Neuropteroid species the ‘‘only possible sternal element is the very narrow area between the sternal apophyseal pits and perhaps along the apex of the internal ridge formed by the meeting of the subcoxae’’ (5 subcoxal sclerites). He interpreted a basisternal sclerite, an area between the sternal apophyseal pits, as a purely secondary development ( Ferris, 1940b: 88). (However, see Matsuda, 1970, discussed below, who disagreed that the sternum was secondarily developed.) Ferris and his students regarded the sclerites on each side of the discrimen to be preepisternum (fig. 19). No one discussed the fate of the laterosternite (5 sternopleurite of Snodgrass, 1935: 163, 192) ( figs. 12, 14 View Figs , 18) until Matsuda (1960: 713) renamed it ‘‘ventropleurite’’ and wrote that it was fused to either the basisternum or the preepisternum or lost. If the sternal apophyseal pits are near the lateral limit of the sternum, the ventropleurite would be laterad of the pits and, although unidentifiable, one wonders why the ventral sclerite is interpreted as only preepisternum rather than ‘‘preepisternum + ventropleurite.’’ However, regardless of whether or not the sternal apophyseal pits mark the lateral edges of the sternum, any sclerites or membranes between them would be sternal because the endosternites are sternal elements. Saether (1980: 14) equated the sternopleurite with the preepisternum, but that interpretation is erroneous because they are separately derived sclerites according to morphologists cited in the preceding paragraphs.
Ferris’s view that the ventral pterothoracic sclerites were of subcoxal origin was embraced by Michener (1944: 172–174) for bees and by Hansen and Cook (1976: 32) and Saether (1980: see preepisternum and mesosternum) for Chironomidae , and probably by other workers for other groups.
Matsuda (1960: 714; 1970) disputed the conclusions of Ferris that the presence of the basisternum was a secondary development by citing evidence that it is present in Thysanura, Ephemeroptera, and most hemimetabolous insects. Matsuda (1970: 21) further challenged the hypothesis of the loss by invagination of the pterosterna by citing a study on the postembryonic development of the thorax of a species of Apis that showed the median longitudinal ridge of the basisternum was ‘‘produced by the migration or proliferation of cells from a low midventral ridge… that suggest[s] that the basisternal median longitudinal elevation was not formed by invagination’’ ( Matsuda, 1970: 21). He also cited Gnophomyia tristissima (Tipulidae) as having widely separated coxae and pleurosternal sutures, while at the same time having a median longitudinal suture (5 discriminal line) ( Matsuda, 1960: 722, 723; 1970: 317, fig. 136b). In a dense, complex, demanding monograph, Matsuda (1970: 88–401) discussed or illustrated the thorax of species of 30 orders of insects and for nearly all he interpreted the presence of pterothoracic sterna. Some taxa with the sternal apophyses close together either lacked ( Matsuda, 1970: figs. 118b, 121) or had reduced pterothoracic sterna: for example, Tipula , which had been regarded as lacking pterothoracic sterna by Rees and Ferris (1939), was considered to possess pterosternal sclerites by Matsuda (fig. 136a). Ferris’s (1940a, 1940b) hypothesis was based on four pterygote orders of insects, but was challenged by Matsuda (1970) based on data for many more orders, both apterygote and pterygote. However, Matsuda’s objections appear not to have been addressed.
For the Coleoptera, Matsuda (1970: 199– 218) regarded the pterothoracic pleural (subcoxal) sclerites to include an epimeron, anepisternum, katepisternum, preepisternum, and trochantin and the sternal sclerites to include a basisternum (including a presternum, an anterior part of the basisternum; fig. 19, also see fig. 18), and spinasternum (fig. 19); his ventral plate included both sternal and pleural elements. Matsuda (1970: 205–214) revisited Campau’s (1940) study of Chauliognathus pennsylvanicus and renamed many of the structures according to a partial loss of the pterosterna and corrected some sclerites and sutures that Campau misunderstood (fig. 19, compare left and right sides). Naomi’s (1988) study of the pterothorax of the Staphylinidae largely followed Matsuda’s interpretation.
More recently, Lawrence et al. (2011: 33– 34), without explanation, cited and endorsed Ferris’s (1940b) hypothesis that the entire ventral and lateral surfaces of the metathorax were pleural sclerites, and adopted metaventrite for the ventral sclerite and, perhaps, so interpreted the mesoventrite, but that is unclear (see Lawrence et al., 2011: 30, char. 159). After the work of Ferris, Lawrence and associates were the next to apply this hypothesis broadly to the Coleoptera .
Matsuda’s work (1960, 1963, 1970) offers a far more complex and nuanced understanding of the pterothoracic ventral sclerites than do the views of Ferris. Matsuda agreed with Weber and Ferris that invagination occurred, but thought only part of the pterosternal sclerites were invaginated in most cases and that sternal loss was secondary. Based on the numerous examples in his 1970 work, insects have wide to narrow pterosterna, depending on the distance between the bases of the apophyses, and some may have no sternal elements at all. However, searching his 1970 monograph, one is hard-pressed to find examples of insects that have no sternal elements at all. His view might be summarized in a statement he made about beetles: ‘‘The degree of invagination of the basisternum and possibly of the preepisternum must vary in different beetles; in some beetles the original basisternal area is presumably present indistinguishably along the median longitudinal suture’’ ( Matsuda, 1970: 214; fig. 19). Because Matsuda’s ideas have not been refuted, the position adopted in the present work is his subtler perception of the evolution of the pterothoracic ventral sclerites. Despite the fact that ‘‘mesoventrite’’ and ‘‘metaventrite’’ were coined to refer to the pleural sclerites that replaced pterothoracic sterna lost through invagination, they are also useful terms to refer to the ventral sclerites that are a composite of sternal and pleural elements.
Mesothorax of Oedichirus: For the mesothorax of Oedichirus geniculatus (figs. 20, 21), the smaller and more posterior of the two lateral sclerites is the mesepimeron separated by the mesopleural suture from the larger mesanepisternum that is distinguished from the mesoventrite by a ridge, the mesanapleural suture. The mesanapleural suture terminates at the mesothoracic constriction near the anterior third of the segment. The mesoventrite lacks a discriminal line. The surface from the anterior margin to the mesothoracic constriction can be considered the mesobasisternum + mesopreepisternum + mesanepisternum, and behind the mesothoracic constriction the median surface is the mesobasisternum + mesopreepisternum. The mesoventrite between and posterior to the mesosternal apophyseal pits is the mesofurcasternum (5 mesosternellum). The mesoventropleurite is theoretically indistinguishably fused between the mesobasisternum and mesopreepisternum, so a remnant of the ventropleurite may still be present. The mesoventrite is separated from the metaventrite by a sinuous mesometaventral junction.
The mesopleura and mesoventrite are coarsely punctate (figs. 20, 21). The mesepimeron has a few coarse punctures along the anterior margin and the mesanepisternum has a cluster of coarse ones in the central region; the moderately long setae of both sclerites are inserted at the anterior margin of the punctures. Anterior to the mesothoracic constriction the punctures have minute setae (fig. 20, inset), whereas the punctures posterior to the constriction have long setae.
Metathorax of Oedichirus : The lateral sclerites of the metathorax (fig. 21) are the long, wide metepimeron dorsally separated by a metapleural suture from the more ventral, much narrowed metanepisternum. Distinguished ventrally from the metanepisternum by the metanapleural suture is the large metabasisternum + metapreepisternum (5 metaventrite) which occupies the ventral surface. The metaventrite is not marked internally by a midlongitudinal ridge nor externally by a discriminal line (5 median longitudinal suture, fig. 19), although the short, shallow, midlongitudinal groove between the metacoxae might be a remnant of the discriminal line (fig. 21). Along the posterior margin of the preepisternum, separated by the metakatepisternal suture, is a narrow metakatepisternum.
Most of the metaventrite is coarsely punctate and near the anterior edge of each puncture is a long seta. The mesocoxal cavity has strong microsculpturing. The mesocoxal cavity is formed by the inflexed posterior portion of the mesobasisternum + mesopreepisternum and the mesofurcasternum and is separated by the mesometaventral junction from the broadly depressed anterior portion of the metabasisternum + metapreepisternum. The mesocoxal cavity is margined posteriorly by a pericoxal ridge (figs. 20, 21) and has a midlongitudinal carina/ridge between the coxae; the anterior portion, on the mesoventrite, is a short intercoxal carina extending from the mesoventral process that continues on the metaventrite as a low, rounded ridge that reaches the pericoxal ridge that borders the posterior margin of the acetabulum.
In common with most Paederinae , the separated mesendosternite is a pair of straight, slender apophyses and a forked metendosternite (fig. 22).
LEGS: The forelegs of Oedichirus , in contrast to the middle and hind legs, are highly modified. The procoxa is elongate, the femur has a long slit ventrally that can open widely. The tibia has multiple rows of diagonally transverse combs on the ventral surface, which is not depressed; the apex is beveled. The basal four tarsomeres are inflated and are discussed in the description above. The function of the femoral opening is unknown, but it may be a secretory region ( Herman, 2010: 17). Detailed examination of the enlarged protarsomeres requires SEM images, but most of the species were represented by so few specimens that exploration of the protarsomeres was impossible. The tibiae of all the legs lack spiniform setae along the length. The metatibial apical region is expanded and flattened and both sides of the apical margin have a comb of closely spaced, slender, spiniform setae. Mesotarsomere and metatarsomere 1 are longer than any of the four distal tarsomeres. The fifth tarsomere of the middle and hind legs originates at the base of tarsomere 4, which extends beneath 5 and which is clothed ventrally by a dense pad of modified setae.
ABDOMEN: A term widely used by taxonomists of staphylinids for the one or two small sclerites between the tergum and sternum is paratergite. The term suggests that the sclerite is beside the tergite, but since the large, median, dorsal sclerite of each segment is the tergum ( Snodgrass, 1935: 76– 78, 82) and not tergite, then the logical name for the adjacent sclerite(s) is paratergum (paraterga) (signifying ‘‘beside the tergum’’). The term is purely descriptive and easily locates the sclerite without suggesting its origin. However, in the Staphylinidae these sclerites are probably of sternal origin ( Herman, 1970: 350; Hansen, 1997: 26; Lawrence et al., 2011: 44, char. 279 [these authors appear to consider paratergites of staphylinids and the laterosternites and parasternites of other Coleoptera to be homologous]; Naomi, personal commun.) and it is possible that in the Staphylinidae they will be renamed to reflect this hypothesis, but until then paratergite is used.
The abdomen of Oedichirus is coarsely and densely to sparsely punctate; the punctures have a seta near the anterior margin. For most species the punctation is more or less evenly distributed, but in some species the punctures are arranged in transverse rows on the apical half of the segment, and most, perhaps all, species have a row of small, setate punctures adjacent to the posterior margin. Punctures of the basal four segments (III to VI) are coarser, larger, and the distribution denser than on VII and VIII. Segments III to VII of Oedichirus have a dorsal and ventral pair of oval, opaquely membranous ‘‘windows’’ in the intersegmental membrane adjacent to the anterior margin (fig. 24). No glands nor glandular reservoirs associated with these structures have been found. Sternum IV of species of some paederine genera have an associated glandular reservoir; species of Oedichirus do not.
Sternum I, often represented in the Paederinae by a narrow, frequently medially divided sclerite near the anterior margin of sternum II, is absent in Oedichirus . Sternum II is as wide as segment III, is fused to the anterior margin of sternum III, has a row of setae along the posterior margin, and a median intercoxal ridge; the anterior margin is bisinuate and lobed medially. Sternum III has a median intercoxal carina that ends in an acute, ventroposteriorly directed point near the middle of the segment. A short, sublateral, posteriorly directed carina extends from the transverse basal ridge. The transverse basal ridge of tergum III has a similar, but shorter, sublateral, posteriorly directed ridge that is usually reduced to a trianguloid projection which herein is referred to as a sublateral point. Three New World species, O. bullaglaber , O. bullahirtus , and O. echinatus , have a third, similar medial projection, the median point (similar to sublateral point of fig. 23 and to that on tergum VIII, fig. 26). Tergum VIII also has a median point in O. exilis , O. clavolateralis , O. bullaglaber , O. bullahirtus , O. echinatus , and O. geniculatus , whereas other species lack it. The median point of tergum VIII is apically open (fig. 114) or closed (fig. 73).
The abdomen of Oedichirus is cylindrical, most segments lack paratergites, and the tergum and sternum of most segments are fused. Segment III for some species has one paratergite (on each side) that is about as long as the segment, narrow, and tapered posteriorly; for these species the tergum and sternum are separated. Paratergite III is present in six New World species ( O. batillus , O. bicristatus , O. hamatus , O. isthmus , O. optatus , and O. sinuosus ). For the other species a carina replaces the paratergite. This carina, the paratergal carina, although small- er, is of similar form and occupies the same position as would the paratergite and extends to, or slightly to well beyond, the spiracle (fig. 23). There may be a feeble line, a remnant of the tergosternal suture, extending posteriorly from the end of the paratergal carina (fig. 23), but the tergum and sternum are fused. Because the paratergal carina and the paratergite are of similar size, form, and position, it seems likely that the former is derived from the latter. Segments IV to VII lack paratergites, but the lateral surface of some segments of some species have a faint ridge just ventrad of the spiracle that may be a remnant of the tergosternal suture. Although segment VII lacks this feeble ridge, the separation of the tergum and sternum is marked posteriorly by an incision that may be the remainder of a tergosternal suture (fig. 25). This incision is tiny in some species, for example O. apiculus , O. glabrihamus , and O. misionesiensis . Tergum and sternum VIII are separated and the segment always lacks paratergites (fig. 26). As is the case for many paederines, sternum VIII of Oedichirus has species-diagnostic features in males, but not in females.
Segment IX differs from the preceding segments in that the base of tergum IX extends from the dorsal (figs. 27, 140) to the ventral side (fig. 28) and the lateroapical corners of the tergum are prolonged posteriorly beyond the median posterior margin and referred to as lateroapical processes (figs. 28, 29). In some paederines tergum IX is divided dorsally, and the halves can be referred to as hemitergites or laterotergites, but for Oedichirus tergum IX is fused middorsally (figs. 27, 140). Tergum IX of Oedichirus differs between males and females. Most males have a dorsoventrally flattened process extending anteriorly from the ventromedial edge of the anterior margin of the tergum; the process is short, wide, and apically rounded to long, moderately wide, and apically rounded to truncate; the females lack this process (fig. 29). The size and form of the anteroventral process of the males varies among species. For males and for most females, the anteroventral edges of tergum IX are separated midventrally (figs. 28, 29), but near the left margin in O. optatus (fig. 195). However, for females of O. dilophus , O. distortus , O. lunatus , O. ohausi , and O. procerus , the anteroventral angles are fused midventrally to form a narrow to moderately wide bridge or strap (figs. 108, 116, 169, 186, 197). The lateroapical processes are long, slender, tapered, apically acute, tubular, ventrally curved, and usually widely separated. In most species the ventral curvature of the lateroapical processes is slight (fig. 158) to moderate and the processes parallel to each other or slightly divergent. Those of O. lunatus are semicircularly curved in lateral view (fig. 168) and widely separated apically in dorsal view (fig. 170). In dried specimens the lateroapical processes nearly touch distad of the base and perhaps do so in live individuals also or perhaps the processes touch only as an artifact of drying. Sternum IX, the median ventral sclerite, of the male is long, wide, and symmetrical or asymmetrical (figs. 28, 37, 45, 109, 117). The more complex female ventral sclerite of IX is discussed in the following paragraph. Tergum X (fig. 27) is embedded in the gap between the lateroapical processes of tergum IX and attached to the middorsal posterior margin of tergum IX. Tergum X is large to small, shield shaped, and has cuticular processes on the posterior margin that is broadly rounded and that may be slightly produced medially. Tergum X of Oedichirus is exposed and only the basal edge is slightly covered by tergum IX.
As briefly discussed already ( Herman, 2010: 26–27), the female genital sclerites exhibit wide variation in the Paederinae . In that same article, these sclerites were illustrated for one species of Oedichirus . Howev- er, despite the large number of species, about 75 % of which were described in the last 40 years ( Fagel, 1971; Lecoq, 1986, 1987, 1990, 1991, 1996; Janák, 1995, 1996, 1998, 2003; Last, 1980; Jarrige, 1978; Hayashi, 1989; Outerelo and Gamarra, 1988), the female genital sclerites of Oedichirus remain unknown. Females of 18 New World species were available for examination. For the species of Oedichirus examined, the median gonocoxal plate is wide, long, lacks styli, and has an associated vulvar plate (figs. 29, 186). The size, form, and presence of the median gonocoxal plate anterior to the vulvar plate varies (figs. 52, 68, 76, 195) and for all species but O. optatus (fig. 195), the median gonocoxal plate is far larger posteriad of the vulvar plate than anteriad (figs. 29, 52). The anterior portion can be large and triangular or trapezoidal (figs. 68, 76, 148), or a narrow strap (figs. 52, 116). For O. lunatus and O. ohausi the anterior portion of the median gonocoxal plate is joined to the fused anteroventral angles of tergum IX (figs. 169, 186). For O. boehmi and O. dilophus , the anterior portion of the median gonocoxal plate is lost and the vulvar plate is attached only to the anterior edge of the posterior portion of the median gonocoxal plate rather than embedded in it (figs. 64, 108).
The vulvar plate of New World species is comprised of an anterior and posterior lobe that, in most species, is embedded in the median gonocoxal plate (figs. 29, 69). The two vulvar lobes are clear and distinct for most species, but for O. dilophus and O. lunatus the demarcation is subtle (figs. 111, 171) and appears to be incomplete. The posterior vulvar lobe is transverse (figs. 69, 118, 159, 185) to more or less rounded (figs. 97, 196) and includes the vulva (figs. 69, 118) and the entrance to the vagina ( Tuxen, 1956: 283); from the posterior vulvar lobe, on the interior side, the spermathecal duct can be seen extending (figs. 169, 171). For most species the anterior vulvar lobe is transverse (figs. 118, 159, 185) or transversely arc shaped (figs. 69, 118, 151) and may wrap around the anterior and lateral margins of the posterior vulvar lobe (figs. 55, 186, 196, 220, 224). For two species, O. boehmi (figs. 64, 67) and O. ohausi (figs. 186, 187), the anterior vulvar lobe envelops the right anterolateral margin of the posterior vulvar lobe, which, for the latter species, is more irregularly shaped than in others. Both vulvar lobes of O. boehmi are strongly sclerotized, unlike other New World species. Generally the two lobes of the vulvar plate are membranous and adorned with variously distributed microstructures, or are without adornment or slightly to strongly sclerotized (fig. 186). The anterior vulvar lobe is smooth or wrinkled (figs. 118, 151, 159), covered with a reticulate fan (fig. 224) or cuticular processes of varying density, form, and length (figs. 55, 69, 111), or is membranous and without adornment (figs. 118, 220). The ornamentation of the posterior lobe includes cuticular processes (fig. 111), cuticular cobble (figs. 69, 88, 196), and reticulation (fig. 159). The cuticular processes may have one to five or six apical fimbriae (figs. 111, 185). The orientation of the vulva, which is often surrounded by membranes and which often appears to be in a depression, is vertical (fig. 69), diagonal (figs. 111, 196), or transverse (fig. 159), and straight (fig. 69) or curved (fig. 118). In the confocal laser scanning images membranous regions that lack surface microstructures are black, with perhaps white lines where the membrane is wrinkled and overlaps itself (fig. 220), and merge with the surrounding black background as though nothing is present. The form of the anterior margin of the median gonocoxal plate and the form, type, and distribution of the adornment of the vulvar plate and its two lobes are diagnostic, perhaps for species or perhaps only for species groups. No general statements are possible about the female genitalia of Old World species because few species were examined.
A sclerotized spermatheca was not found. The spermathecal duct is long and extends from the vulva far into the abdomen (figs. 169, 171).
The aedeagus is asymmetrical and diagnostic for Oedichirus species. The median lobe varies considerably and has many features useful for identification of species (for examples see illustrations herein and those of Fagel, 1971; Lecoq, 1986; and Janák, 2003) and some characters for defining groups of species. The ventral sclerite of two clusters of species has an apicoventral process (figs. 142, 212) that extends from the apical margin and, in dorsal view, curves laterally to the right and the apex is directed anteriorly. The surface and edges of the ventral sclerite have species specific tumescences and spines. The condition of the parameres is partially diagnostic for species, but characters of the median lobe are more easily seen and exhibit more variation. The parameres taper apically from the base, are long, slender, and cylindrical to short, broad, and flat. Short, broad, flat parameres are fused to the lateral sides of the median lobe. Long slender parameres are fused to the median lobe basally. The anterior portion of a basally broad, apically slender paramere is fused to the median lobe. For some species the paramere is so completely fused to the median lobe that only a shallow depression along the margin of an elongate tumescence marks the limit of the paramere. The internal sac is difficult to evert, but has processes, sclerites, spines, and microstructures that may help define species and/or perhaps groups of species. The internal sac of four species, O. bullaglaber , O. geniculatus , O. misionesiensis , and O. speculifrons (figs. 77, 146, 178, 215), everted spontaneously while clearing the aedeagus. The four exhibit significant, perhaps species-diagnostic differences. Two of them ( O. speculifrons and O. misionesiensis ) are in the same species group based on other characters, also share features of the internal sac. The other two species, O. geniculatus (fig. 152) and O. bullaglaber (fig. 78), are in different species groups and their respective internal sacs differ from each other and from the other two species. A wealth of diagnostic and phylogenetic characters may become available if techniques are developed that permit consistent, reliable eversion of the internal sac. Note that even without eversions, spines and processes often can be seen on the internal sac through the cuticle of the aedeagus. However, I have never been able to consistently and adeaquately clear the inside of the aedeagus of miscellaneous tissue to render structures of the internal sac visible. Furthermore, even when visible, the interspecific relationship of the structures is obscure—if the internal sac of one species is visible that of other species often is not.
Keys to New World Species of Oedichirus (Two keys are presented here, one for males and one for females.)
Key 1. Males of Oedichirus (Sternum VIII with emargination of posterior margin and with surface modified by depressions, tumescences, or clusters of spiniform setae. Males are unknown for O. brunneus , O. hamatus , O. lunatus , O. ohausi , O. procerus , and O. sparsipennis )
1. Sternum VIII weakly, barely detectably, and broadly emarginate (fig. 184); surface with subapical comb of spinelike setae; Costa Rica, Nicaragua (?).. neotropicus View in CoL
– Sternum VIII with distinct, shallow to deep emargination (figs. 102, 163); surface with or without comb of spinelike setae (figs. 46, 122).................. 2
2 (1). Sternum VIII with asymmetrical emargination of posterior margin (figs. 73, 85, 102, 114)..................... 3
– Sternum VIII with symmetrical emargination of posterior margin (figs. 46, 53, 59).... 12
3 (2). Tergum VIII with median point extending from transverse basal ridge (fig. 26)... 4
– Tergum VIII without median point extending from transverse basal ridge... 9
4 (3). Tergum III with median point extending from transverse basal ridge (as for sternum VIII, fig. 73)................... 5
– Tergum III without median point extending from transverse basal ridge...... 6
5 (4). Aedeagus with posterior margin of ventral sclerite notched (fig. 71); tumescence of sternum VIII asetate and gradually sloped posteriorly (figs. 73, 74); Santa Catarina , Brazil.................. bullaglaber View in CoL
– Aedeagus with posterior margin of ventral sclerite entire (fig. 81); tumescence of sternum VIII setate and sharply sloped posteriorly (figs. 84, 85); São Paulo, Brazil.................... bullahirtus View in CoL
6 (4). Aedeagus with blunt peg on right lateroapical margin of ventral sclerite dorsad of apicoventral process (figs. 98, 99); Rio de Janeiro, Brazil............. clavulus View in CoL
– Aedeagus not as described above.... 7
7 (6). Aedeagus with broad lobe on left side of posterior margin of ventral sclerite (fig. 90); ventral sclerite with knob extending laterally from base of right margin of apicoventral process (fig. 89); Rio de Janeiro, Brazil......... clavolateralis View in CoL
– Aedeagus not as described above.... 8
8 (7). Aedeagus with flattened lobe extending dorsally from ventral sclerite near base of apicoventral process (fig. 129); labrum without tubercle near base of submedial denticle; Rio de Janeiro, Brazil.... exilis View in CoL
– Aedeagus without adornment near base of apicoventral process (fig. 141); labrum with tubercle near base of submedial denticle (fig. 136); Rio de Janeiro, Brazil..................... geniculatus View in CoL
9 (3) Paratergite of segment III present, paratergal carina absent.............. 10
– Paratergite of segment III absent, paratergal carina present (fig. 23)....... 11
10 (9). Sternum VIII with irregularly arranged, transverse cluster of subapical, spinelike setae (fig. 163); sternum VII with large, median cluster of short spinelike setae (fig. 165); Jalisco, Mexico..... isthmus View in CoL
– Sternum VIII with strongly curved, evenly arranged, transverse row of subapical, spinelike setae (fig. 203); sternum VII with small, median cluster of long, slender spinelike setae (fig. 205); Yucatan, Mexico.................... sinuosus View in CoL
11 (9). Sternum VIII with prominent process on right side of emargination of posterior margin (fig. 114); sternum VII without median cluster of spinelike setae; Ecuador.................... distortus View in CoL
– Sternum VIII without process on posterior emargination (fig. 122); sternum VII with dense median cluster of spinelike setae (fig. 121); Dominican Republic......................... dominicanensis View in CoL
12 (2). Paratergite of segment III present, paratergal carina absent.............. 13
– Paratergite of segment III absent, paratergal carina present (fig. 23)....... 15
13 (12).Sternum VIII with deep, narrow emargination (fig. 193); Pará, Maranhão, Mato Grosso, Brazil............. optatus View in CoL
– Sternum VIII with wide, shallow to moderately deep emargination (figs. 53, 59).......................... 14
14 (13).Sternum VIII without cluster of spinelike setae (fig. 53); emargination of posterior margin moderately deep (fig. 53); Pará, Brazil.................... batillus View in CoL
– Sternum VIII with dense cluster of spinelike setae medially (fig. 59); cluster divided medially and with setae lateroposteriorly directed from middle; emargination of posterior margin shallow (fig. 59); São Paulo, Brazil............ bicristatus View in CoL 15 (12). Tergum III with median point extending from transverse basal ridge (as for sternum VIII, fig. 73); Parana, Brazil...................... echinatus View in CoL
– Tergum III without median point... 16
16 (15). Head with V-shaped depression dorsally; pronotum sparsely punctate and with deep, submedial, punctate groove; elytra longer than conjoint width (EW/ EL 5 0.9–0.8) and dorsal surface concave; Bahia, Brazil...... boehmi View in CoL
– Head without V-shaped depression dorsally; pronotum moderately densely punctate and with submedial cluster of punctures; elytral conjoint width great- er than length (EW/EL 5 1.1–1.4) and dorsal surface flat to convex..... 17
17 (16). Aedeagus with apicoventral process extending to right from apex of ventral sclerite (figs. 42, 212); sternum VIII without subapical, submedian carina (figs. 46, 155); sternum VIII with transverse basal ridge entire, not separated medially (figs. 46, 47).......... 18
– Aedeagus without apicoventral process (fig. 104); sternum VIII with short, subapical, submedian carina adjacent to median depression (fig. 106); sternum VIII with transverse basal ridge separated medially (fig. 107); Bolivia......................... dilophus View in CoL
18 (17). Aedeagus with posteriorly directed, apically truncate, conical peg-boss at base of left side of apicoventral process (figs. 173); Misiones, Argentina..................... misionesiensis View in CoL
– Aedeagus without peg-boss at base of apicoventral process (fig. 42, 212)... 19
19 (18). Aedeagus with apically acute, spiniform process at base of apicoventral process on apical margin of right lateral side of ventral sclerite (figs. 41, 44); Santa Catarina , Brazil.......... apiculus View in CoL
– Aedeagus without spiniform process as above...................... 20
20 (19). Aedeagus with broad flat surface of apicoventral process facing ventrally and nearlyhorizontal (fig. 154); ventral sclerite of aedeagus with small, pointed lobe on right margin near base of apicoventral process (fig. 153); Santa Catarina , Brazil........ glabrihamus View in CoL
– Aedeagus with flat surface of apicoventral process posteriorly facing and nearly vertical (figs. 211, 212); ventral sclerite of aedeagus without pointed lobe on right margin near base of apicoventral process (fig. 212); Santa Catarina , Brazil........ speculifrons View in CoL
Key 2. Females of Oedichirus View in CoL (Sternum VIII unmodified. Females unknown for O. apiculus View in CoL , O. bicristatus View in CoL , O. clavolateralis View in CoL , O. clavulus View in CoL , O. dominicanensis View in CoL , O. echinatus View in CoL , O. exilis View in CoL , O. glabrihamus View in CoL , O. isthmus View in CoL , O. misionesiensis View in CoL , and O. sinuosus View in CoL )
1. Paratergite of segment III present, paratergal carina absent.............. 2
– Paratergite of segment III absent, paratergal carina present (fig. 23)....... 4
2 (1). Median gonocoxal plate strongly asymmetrical (fig. 195) ; tergum IX with right anteroventral angle extending across width of segment to left side and left anteroventral angle absent (fig. 195); legs unicolored, without dark femorotibial spot (as in fig. 1); Pará, Maranhão, Mato Grosso, Brazil............. optatus View in CoL
– Median gonocoxal plate symmetrical (figs. 52, 157); tergum IX with left and right anteroventral angles of approximately equal size (figs. 52, 157); legs bicolored (as in fig. 2) or unicolored (as in fig. 1)..... 3
3 (2). Tergum IX with large, posteriorly direct- ed, flat spur on ventrolateral edge below lateroapical process (figs. 157, 158); legs bicolored, with dark femorotibial spot (as in fig. 2); Peru............ hamatus View in CoL
– Tergum IX without spiniform process on ventrolateral edge (fig. 52); legs unicolored, without dark femorotibial spot (as in fig. 1); Pará, Brazil........ batillus View in CoL
4 (1). Tergum III with median point (as for sternum VIII, fig. 73)............ 5
– Tergum III without median point.... 6
5 (4). Body bicolored, head and pronotum dark reddish brown to black, elytra and abdomen pale to dark reddish brown; Santa Catarina , Brazil.......... bullaglaber View in CoL
– Body unicolored dark reddish brown; São Paulo, Brazil............ bullahirtus View in CoL
6 (4). Tergum VIII with median point (fig. 26)......................... 7
– Tergum VIII without median point..... 9
7 (6). Tergum IX with long, semicircularly curved, lateroapical process (figs. 168, 170); anteroventral angles fused to each other and to tergum IX (fig. 169); Bahia, Brazil................... lunatus View in CoL
– Tergum IX with moderately long, slightly curved lateroapical process (as in fig. 158); anteroventral angles separated from each other and from tergum IX (fig. 68)... 8
8 (7). Median gonocoxal plate anteriad of vulvar plate triangular with strongly sloping lateral sides (fig. 68); lateroapical process of tergum IX less than two times longer than midbasal length of tergum (LLaP/T9 5 2.0); São Paulo, Brazil...... brunneus View in CoL
– Median gonocoxal plate anteriad of vulvar plate triangular to trapezoidal and with gradually sloping lateral sides (figs. 148–150); lateroapical process of tergum IX two or more times longer than midbasal length of tergum (LLaP/ T9 5 2.0–3.0); Rio de Janeiro, Brazil................... geniculatus View in CoL
– Median gonocoxal plate anteriad of vulvar plate trapezoidal with gradually sloping lateral sides (fig. 206); lateroapical process of tergum IX about two times longer than midbasal length of tergum (LLaP/T9 5 1.7); Santa Catarina , Brazil............ sparsipennis
9 (6). Tergum IX with anteroventral angles fused medially (figs. 108, 116)..... 10
– Tergum IX with anteroventral angles separated medially (figs. 29)...... 13
10 (9). Vulvar plate densely covered with apically acute cuticular processes (fig. 110); median gonocoxal plate absent anteriad of vulvar plate (fig. 108); Bolivia........................ dilophus
– Vulvar plate without apically acute, cuticular processes (fig. 118); median gonocoxal plate anteriad of vulvar plate narrow and straplike (figs. 116, 197)....... 11
11 (10). Labrum with 2 denticles; elytral disc concave; Bahia, Brazil..... procerus
– Labrum with 4 denticles; elytral disc concave or flat................ 12
12 (11). Legs bicolored, with dark femorotibial spot (as in fig. 2); anterior vulvar plate not sclerotized (fig. 118); tergum VIII with broadly emarginate posterior margin and with small median lobe; Ecuador................... distortus
– Legs unicolored, without dark femorotibial spot (as in fig. 1); anterior vulvar plate partially sclerotized (fig. 187); tergum VIII with broadly rounded posterior margin and without distinct median lobe; São Paulo, Brazil....... ohausi
13 (9). Head with V-shaped depression dorsally; median gonocoxal plate absent anteriad of vulvar plate (fig. 64); Bahia, Brazil.................. boehmi
– Head without V-shaped depression dorsally; median gonocoxal plate present anteriad of vulvar plate (figs. 183, 210)....................... 14
14 (13). Legs bicolored, with dark femorotibial spot (as in fig. 2); tergum VIII with broadly emarginate posterior margin; Costa Rica, Nicaragua... neotropicus
– Legs unicolored, without dark femorotibial spot (as in fig. 1); tergum VIII with shallowly emarginate to rounded posterior margin; Santa Catarina , Brazil................ speculifrons
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Oedichirus Erichson
| Herman, Lee H. 2013 |
Elytrobaeus
| Sahlberg, R. F. 1847: 801 |
Oedichirus
| Herman, L. H. 2010: 36 |
| Herman, L. H. 2010: 36 |
| Navarrete-Heredia, J. L. & A. F. Newton & M. K. Thayer & J. S. Ashe & D. S. Chandler 2002: 293 |
| Blackwelder, R. E. 1952: 269 |
| Blackwelder, R. E. 1944: 131 |
| Sharp, D. S. 1876: 338 |
| Erichson, W. F. 1840: 685 |
| Erichson, W. F. 1840: 685 |
| Erichson, W. F. 1840: 684 |
| Erichson, W. F. 1839: 29 |