
Adalaria slavi, Martynov, Alexander, Korshunova, Tatiana, Sanamyan, Nadezhda & Sanamyan, Karen, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.188931 |
|
publication LSID |
lsid:zoobank.org:pub:179F84F7-CB72-4AB7-877A-BAD332E57FEE |
|
DOI |
https://doi.org/10.5281/zenodo.5694800 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD87DF-4019-FFA1-FF51-F927FED0CEBB |
|
treatment provided by |
Plazi |
|
scientific name |
Adalaria slavi |
| status |
sp. nov. |
Adalaria slavi View in CoL sp. nov.
( Figures 3 View FIGURE 3 C, G; 6D–F; 7B; 9; 11A–D; 12A, B. Table 2)
Type Material. Holotype, ZMMU Lc-37456, (23 mm length), NW Pacific near Kamchatka peninsula, Starichkov Id., 20–26 m, collected by T.A. Korshunova and A.V. Martynov, 14.08.2008. Paratypes, ZMMU Lc-37457, 28 specimens, same locality and collectors as holotype 19.08.2008. Paratypes, ZMMU Lc-37458, seven specimens (one dissected) same locality and collectors as holotype, at 18–24 m depth, large boulders, 19.08.2008. Paratypes, ZMMU Lc-37459, five specimens (three dissected), same locality and collectors as holotype, 14.08.2008. Paratypes, ZMMU Lc-37460, 10 specimens, same locality and collectors as holotype, 19.08.2008. Paratypes, ZMMU Lc-37461, 9 specimens, same locality and collectors as holotype, 19.08.2008. Paratype, ZMMU Lc-37462, one dissected specimen, same locality and collectors as holotype, 19.08.2008.
Type locality. NW Pacific, SE Kamchatka, Starichkov Id., 18– 26 m.
Etymology. This species is named in honour of Vyacheslav G. Shipilov, captain of the boat “Chaika” in recognition of his generous help in organizing scuba diving.
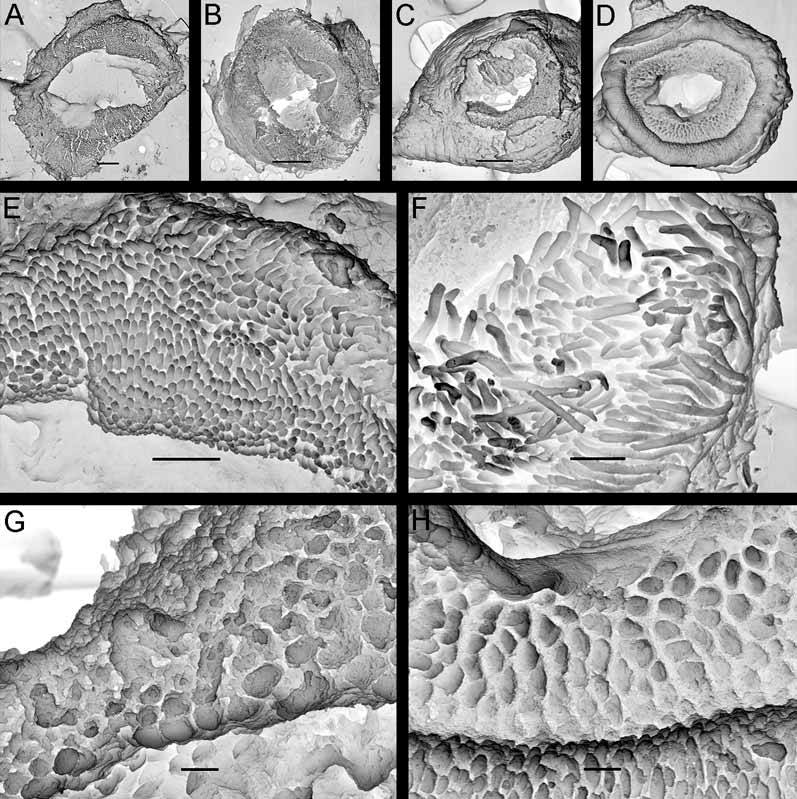
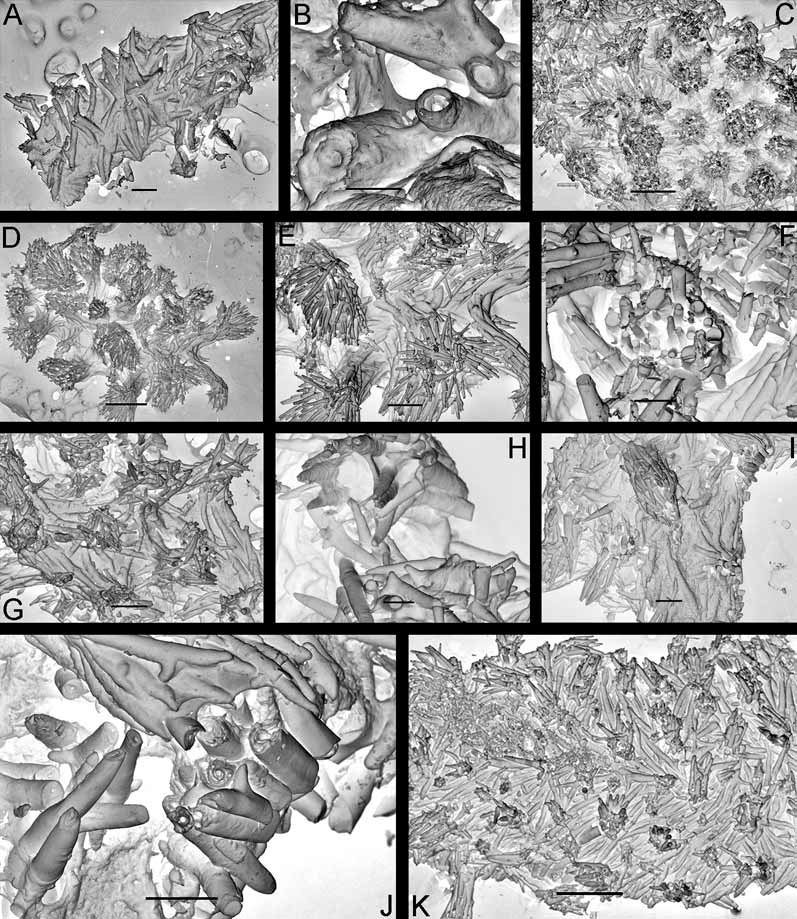
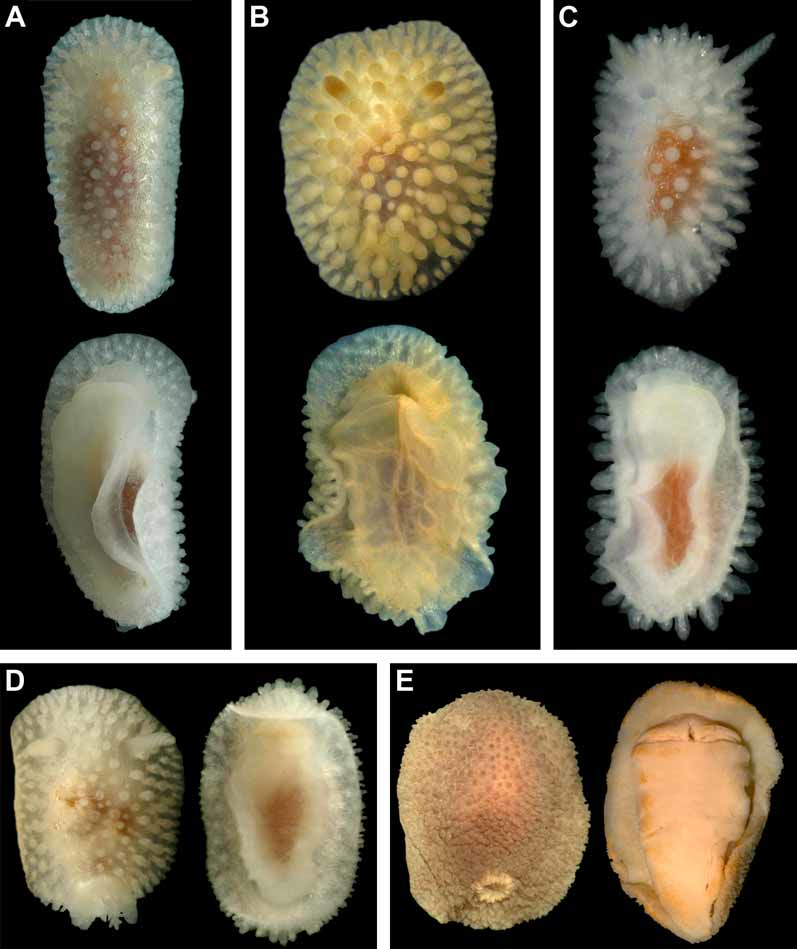
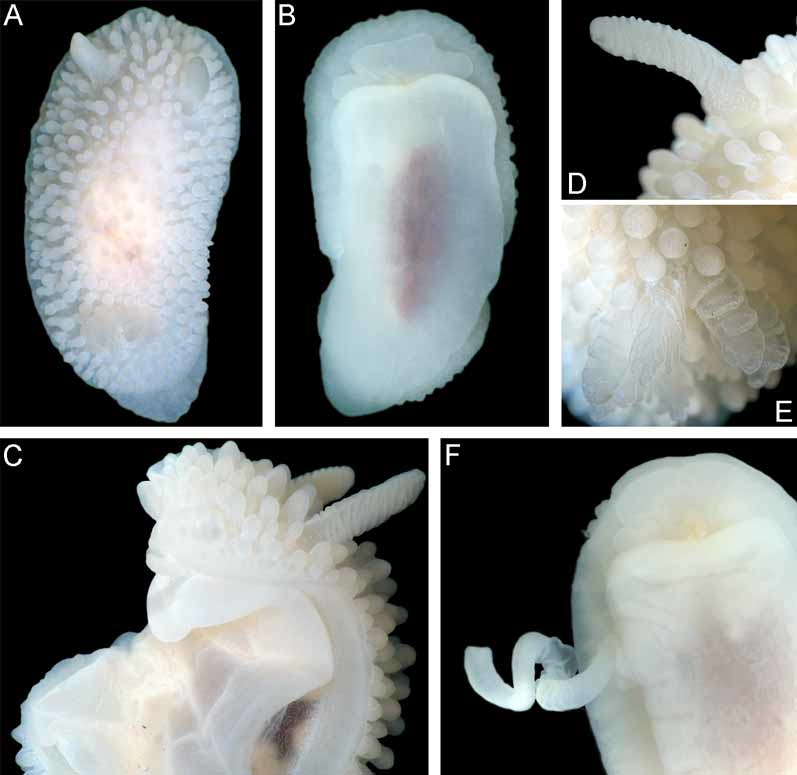
Description. External morphology. The length of the holotype is 23 mm and width 11.5 mm ( Fig. 9 View FIGURE 9 A–B). The length of 20 living specimens ranged from 8 to 23 mm, the width ranged from 4.5 to 11.5 mm. The consistency of living animals is rather soft. The notum is moderately broad, rounded in front and posteriorly. The rhinophores are long and retracted into sheaths with smooth edges, except for 5–6 tubercles of various size that are connected with edge of each sheath ( Fig. 9 View FIGURE 9 D). The rhinophoral sheath edges are capable of some contraction in living specimens. There are 11–14 rhinophoral lamellae. The clavus of the rhinophore has a low ridge posteriorly. The notum is densely covered with inflated cylindrical or almost globular tubercles on a short stalk. Tubercles in the central notal area are somewhat wider and more globular than those at the notal edge. Larger tubercles are regularly intermingled with smaller ones. The rays of spicules radiating from the bases of tubercles form a sort of network under the surface of the apparently soft notum ( Figs. 6 View FIGURE 6 D–E). The spicules are not conspicuous externally. Each tubercle contains dense bundles of spicules, not protruding through the tubercle surface. The strongly calcified spicules are of various sizes, most of them are solid ( Fig. 6 View FIGURE 6 F). The gill cavity is absent. Six to eight bi- and tripinnate gills form an almost complete semicircle around the anus, and one tubercle may be present just behind the anus. ( Fig. 9 View FIGURE 9 E). Three gills were detected in a juvenile of 5–6 mm length. The oral veil is large, since it consists of two pairs of processes: a single, broad trapezoid upper triangular projection that is not medially fused with the hyponotum, and two flattened lobes below ( Fig. 9 View FIGURE 9 B). The foot is broad, anteriorly rounded, and posteriorly slightly projecting beyond the notum in crawling animals forming a rounded tail ( Fig. 9 View FIGURE 9 A).
Colour. The living specimens are milky white, slightly transparent, with a brownish intestine scarcely visible in the middle of the notum. The integument of the notum (including hyponotum) and rhinophores are densely covered with small, faint opaque white dots ( Figs. 9 View FIGURE 9 A, C). On the tubercle tops and rhinophoral lamellae edges the white pigment is almost entirely absent. The gill edges are covered with white dots. The white gonad shines through mature animals (including the holotype), and ventrally the reddish digestive gland can be seen.
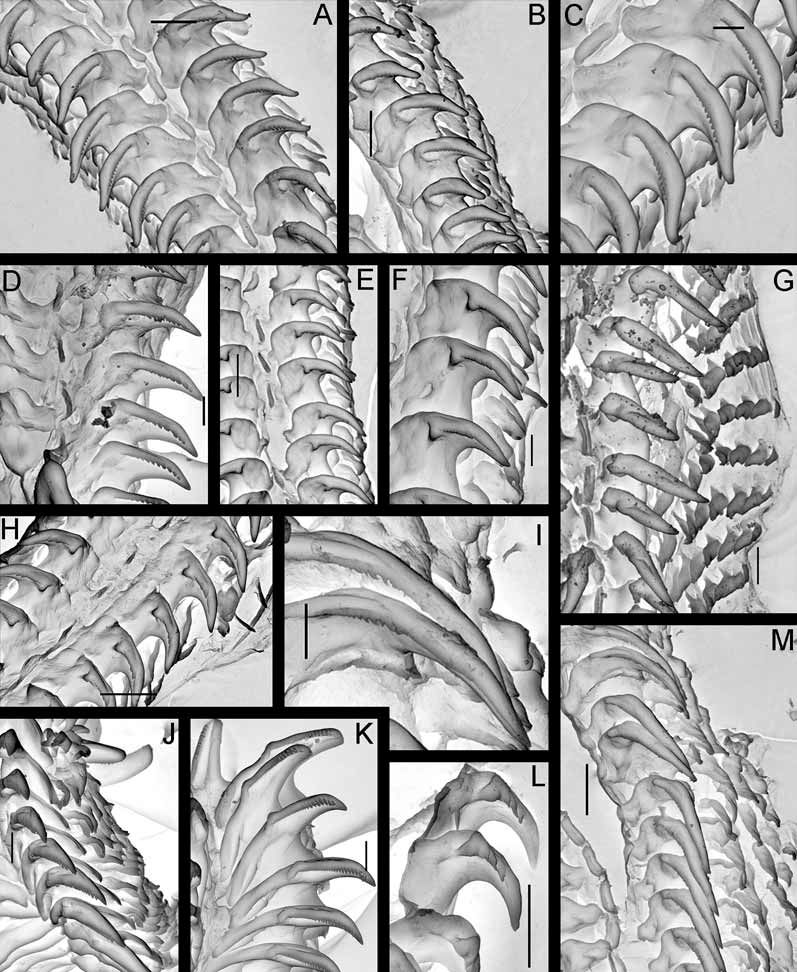
Anatomy. Digestive system. The anterior part of the buccal bulb is modified into the prominent, sessile buccal pump ( Fig. 7 View FIGURE 7 B). The buccal pump is fully banded by a relatively narrow peripheral muscle ( Fig. 7 View FIGURE 7 B). The lateral sides of the buccal pump are provided with thin muscular fibres. The salivary glands are massive triangular lobes ( Fig. 7 View FIGURE 7 B, left figure). The rounded labial disk is covered by yellowish cuticle bearing fine, knob-like labial elements ( Figs. 3 View FIGURE 3 C, G). The radular formula in six studied specimens (15–21 mm length) is 27– 32 x 6–9.1.1.1.9–6. Radular teeth are slightly yellowish. The central tooth is small, elongated, rectangular and folded ( Figs. 11 View FIGURE 11 A, D). The first lateral tooth is provided with a long, wide base and a strong slightly curved beak-shaped cusp, bearing 10–15 small denticles ( Fig. 11 View FIGURE 11 C). The outer denticles gradually reduce in size towards the internal ones. Outer lateral teeth have slightly elongated bases, with a curved, hooked cusp on its lateral corner; all are similar in size and shape ( Figs. 11 View FIGURE 11 A, C). In smaller specimens (8–9 mm length) the denticles of the innermost lateral teeth are relatively larger and fewer (8–10) ( Fig. 11 View FIGURE 11 D) and the cusp itself is straighter. This condition is somewhat similar to the condition in juveniles of Adalaria proxima ( Fig. 11 View FIGURE 11 L) and adult specimens of Adalaria olgae sp. nov. ( Figs. 11 View FIGURE 11 F, H). The stomach is relatively small and narrow. A stomach caecum is absent.
Circulatory system. In the pericardial sac a triangular posterior auricle and a smaller sized oval ventricle are present. The blood gland is rather large in relation to the central nervous system, lies above it and comprises from both posterior and anterior lobes.
Central nervous system. The cerebral and pleural ganglia are well separated, the latter being somewhat larger in size. The optic nerve is very short. The eyes are not large, with black pigment in all studied specimens. The pedal ganglia are similar in size to the cerebrals, lay below them and are connected to them by very short connectives. The rhinophoral ganglia are spherical. The buccal ganglia are slightly oval. Gastroesophageal ganglia are not differentiated. Six pairs of cerebral nerves, three pleural and three pedal ones are detected.
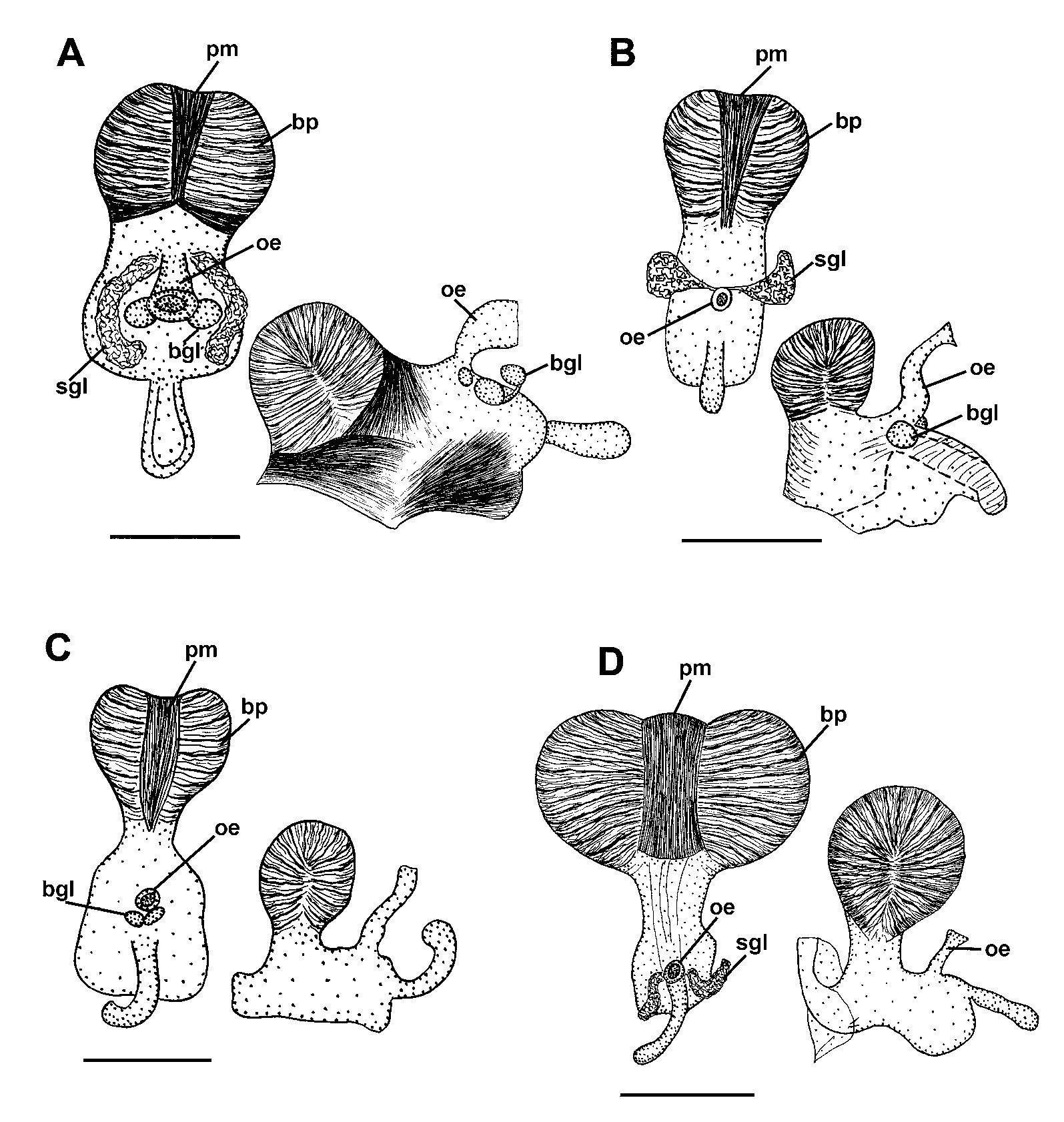
Reproductive system. ( Figs. 12 View FIGURE 12 A, B). The ampulla is moderately short and narrow ( Figs. 12 View FIGURE 12 A, B, a). The post-ampullar duct bifurcates into a long vas deferens and a short proximal oviduct ( Fig. 12 View FIGURE 12 B, pr and pov). The prostate has two distinct parts; a proximal, narrow, rather long convoluted duct partially encircles the bursa copulatrix, a distal, short but greatly swollen part is wrapped within a thin sheath and forms a few lobes ( Fig. 12 View FIGURE 12 A, pr). The prostate transits to a long single-looped penial sheath, which contains several loops of the ejaculatory duct ( Fig. 12 View FIGURE 12 A, psh). The inverted penial sheath and the ejaculatory duct (penis) is long and rather thick, without spines and additional terminal processes ( Fig. 9 View FIGURE 9 F). The moderately sized, globular bursa copulatrix contains some pinkish-red substance; it enters into the vagina via a short narrow stalk ( Fig. 12 View FIGURE 12 B, bc). The proximal oviduct ( Fig. 12 View FIGURE 12 B, pov) is short and rather straight; it extends from near to the junction of ampulla and prostate to the vagina at the bases of the seminal receptacle and bursal stalk. The seminal receptacle is wide, swollen, similar in diameter to the vagina and appears as its prolongation rather than as a separate structure ( Fig. 12 View FIGURE 12 B, rs). The vagina is a long, wide and convoluted duct ( Fig. 12 View FIGURE 12 A, v); near its opening, it has an additional pouch, the vaginal bursa ( Fig. 12 View FIGURE 12 A, vb), and then it opens via a short distal descending part ( Fig. 12 View FIGURE 12 A, pv) and also transits to the off-white nidamental glands by a short wide indistinct distal oviduct ( Fig 12 View FIGURE 12 A, dov).
Biology. Specimens were found predominantly on large boulders covered with several species of encrusting bryozoa, at 18–26 m depth, where it is a very common species.
Distribution. Presently known only from the type locality.
Remarks. Adalaria slavi sp. nov. is well distinguished from other species of the genus by a number of characters. The present species is similar to A. proxima ( Fig. 11 View FIGURE 11 G) and A. loveni in the number of outer laterals (up to 9) but markedly differs regarding the shape of the first lateral tooth (beak shaped covered with small denticles instead of smooth straight cusp), shape of the prostate comprising two parts, different shape of the notal tubercles and characteristic opaque white small dots densely scattered all over the dorsal body side. Adalaria tschuktschica Krause, 1885 ( Figs. 11 View FIGURE 11 I, M) and the poorly described Lamellidoris spiculoides Volodchenko, 1941 were considered as nomina dubia by Martynov (2005) and Millen (2006); they differ from Adalaria slavi in having spiniform elongated notal tubercles, by the shape of the first lateral teeth and by a considerably fewer number of outer lateral teeth (5–6 instead of 6–9). The recently described NE Pacific A. evincta Millen, 2006 significantly differs from Adalaria slavi by the presence of globular tubercles on a very narrow stalk with protruding long spicules, by the differently shaped lateral teeth, the smaller number of outer laterals (3–6 instead of 6–9), the long, convoluted, narrow prostate, and by the colouration. Specimens of Adalaria jannae Millen, 1987 , have been found in course of the present study from Kamchatka waters, but shallowly and never together with the new species ( Fig. 8 View FIGURE 8 A). They are well distinguished externally from Adalaria slavi in having smaller and more slender notal tubercles, a very hard dorsal notum with a strong network of spicules shining through, the presence of a well defined postbranchial gland, and the semitransparent white or yellowish colour without opaque white dots. Internally A. jannae ( Figs. 11 View FIGURE 11 J, K) also clearly differs from Adalaria slavi by its radula that is entirely devoid of central teeth, sharper denticles on the first lateral tooth cusp, fewer outer laterals (4–6 instead of 6–9), and a shorter ejaculatory duct in the reproductive system. Finally, the present species markedly differs from the sympatric Adalaria olgae sp. nov., which inhabits the same depth, by its white colour, bi- and tripinnate gills instead of unipinnate ones, distinct tentacle lobes on the oral veil, a sessile buccal pump, the shape of the first lateral teeth, and by a larger number of outer lateral teeth which differ in their shape. Adalaria slavi is readily distinguished from all known Adalaria species by having a large trilobed oral veil with paired lower lobes and an entire upper lobe, in combination with other features such as body size, colour, and radular features that are summarized in Table 2.
The present species, like at least most other Adalaria and Onchidoris species (see Millen 1987; Schmekel & Portmann 1982; present study, Figs. 12 View FIGURE 12 B; 15A, C), shows an arrangement of reproductive organs that differs from usual doridoidean systems with oocytes and allosperm entering the female gland mass via separate ducts (i.e., oviduct and uterine duct): the proximal oviduct does not enter the female gland mass but connects to the vaginal system; oocytes and allosperm enter the female gland mass via a distally situated, combined duct. Contrary information on Adalaria jannae , A. proxima (Alder & Hancock, 1854) , Onchidoris bilamellata (L., 1767) and O. muricata ( Müller, 1776) by Fahey & Valdés (2005) likely are due to observational errors (Thompson 1966; Millen, 1987; own reexaminations of A. jannae , O. bilamellata and O. muricata ).

| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |