
Bosminopsis africanus ( Daday, 1908 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5052.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:354F50D5-2AB9-4367-84E5-D1365566197D |
|
DOI |
https://doi.org/10.5281/zenodo.5579000 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC87FF-FFD7-190F-1199-FB6EFA9E907F |
|
treatment provided by |
Plazi |
|
scientific name |
Bosminopsis africanus ( Daday, 1908 ) |
| status |
|
Bosminopsis africanus ( Daday, 1908)
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Africa: Daday 1908: 212 ( Bosminella Anisitsi var. africana ); Daday 1910: 140–141, 152, 154, Pl. 8: Figs 1–2 View FIGURE 1 View FIGURE 2 ( Bosminella Anisitsi var. africana ); Brehm 1913: 36–39, Figs 3–6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ( deitersi ); Dumont & Verheye 1984: 315, Fig. 3 View FIGURE 3 ( deitersi ); Kořínek 1984: 47–48, Figs 22–23 ( deitersi ).
Asia: Brehm 1963: 170–171, Figs 16–19 ( deitersi ); Uéno 1966: 103–104, Figs 31–40 ( deitersi ); Mamaril & Fernando 1978: 133, Figs 98–99 ( deitersi ); Idris 1983: 33, 35, Figs 16 ( deitersi ); Rane 1984a: 668 ( deitersi ), 668– 669, Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ( devendrai ); Rane 1984b: 714 ( deitersi ); Michael & Sharma 1988: 99–100, Fig. 30a–e ( deitersi ); Venkataraman 1988: 232, Fig. 2 View FIGURE 2 ( deitersi ); Venkataraman 1995: 384, Figs 49–52 ( deitersi ); Maiphae, 2014: 34–35, Fig. 19 ( deitersi ).
Type locality. “Nyassa” ( Daday 1908) = Lake Victoria , Tanzania .
Type material. Lost. Absent in the Collectio Dadayana , the Hungarian Natural History Museum, Budapest, Hungary ( Forró & Frey 1982) .
Material examined here. See Table 1 View TABLE 1 .
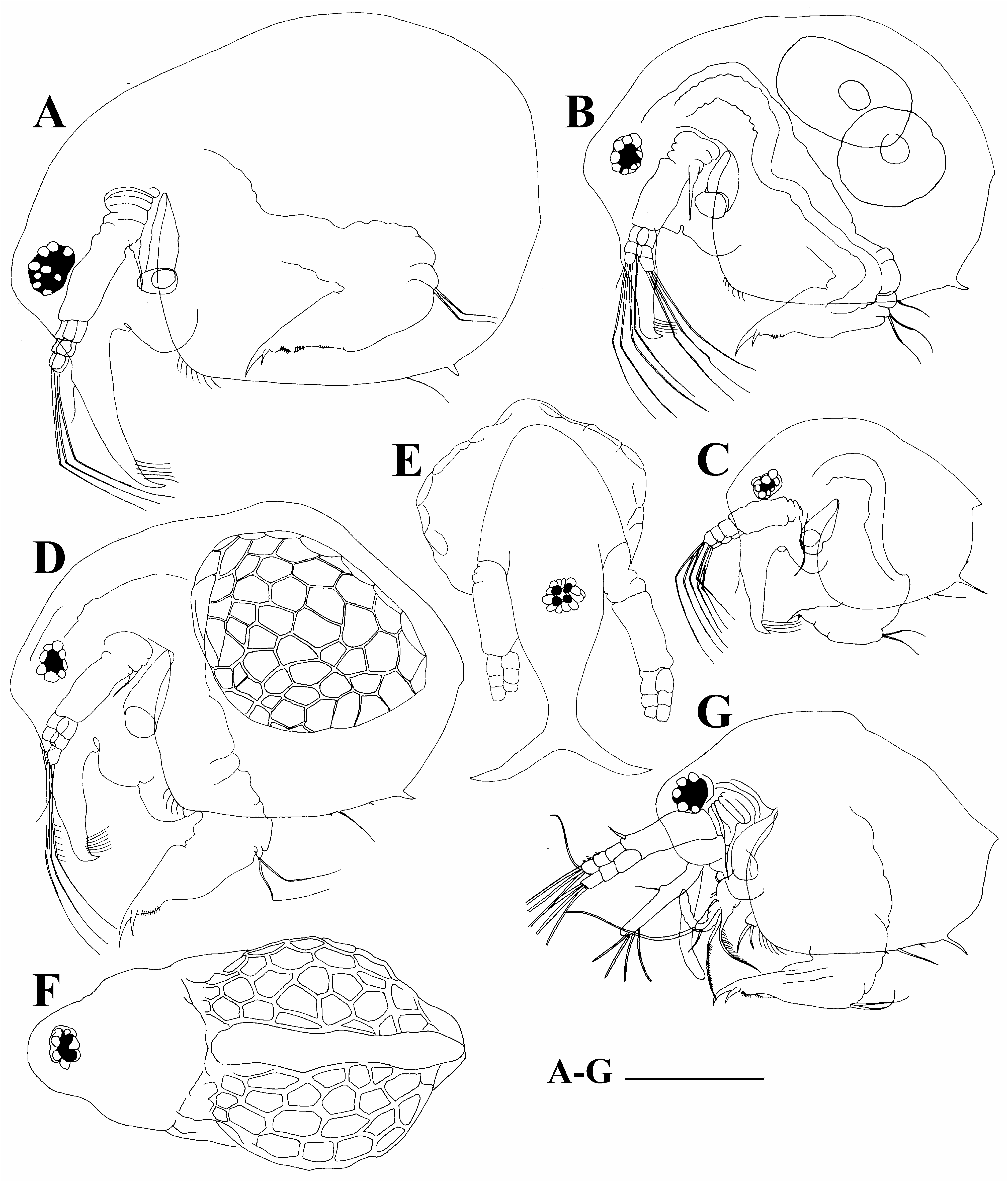
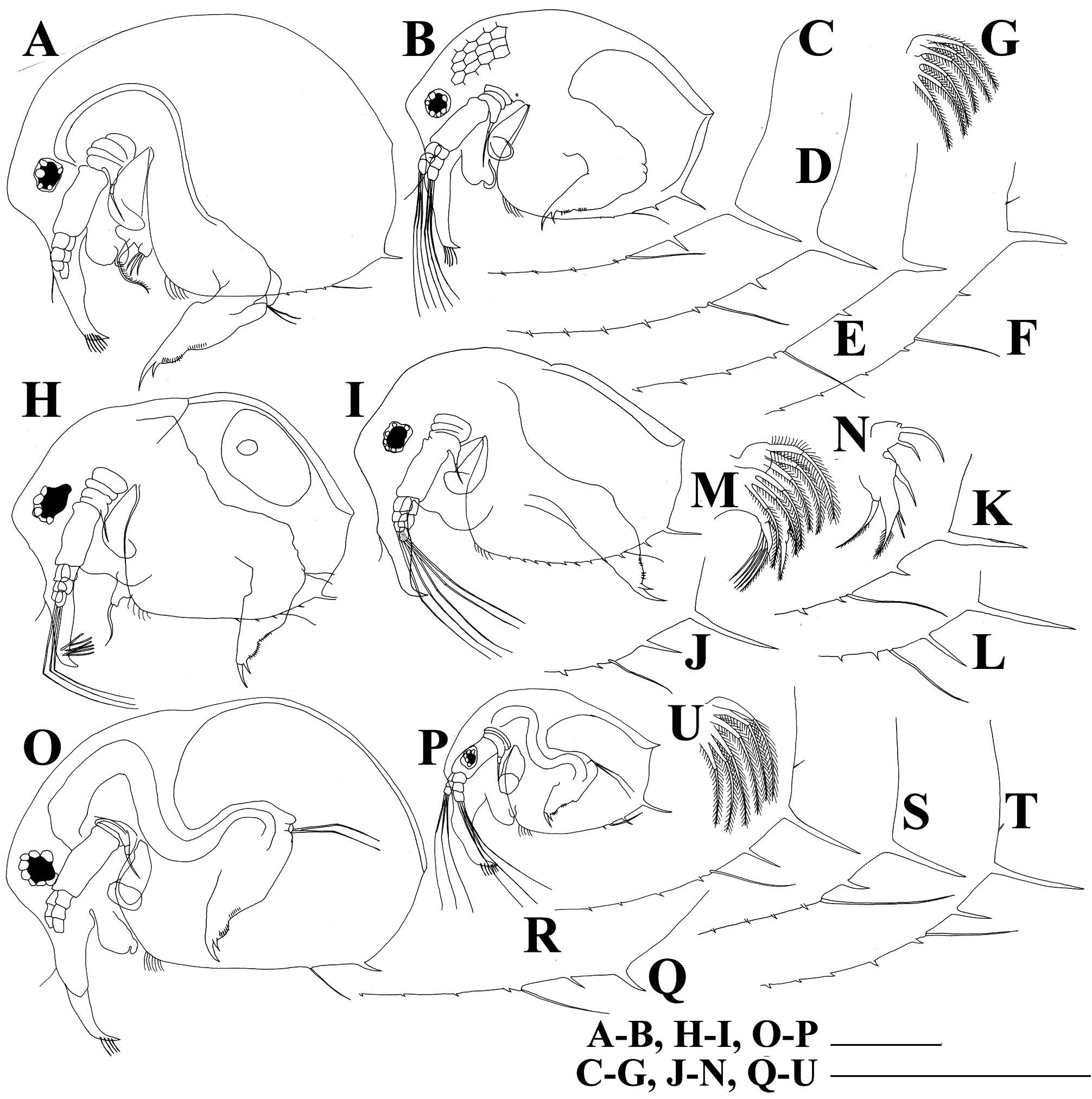
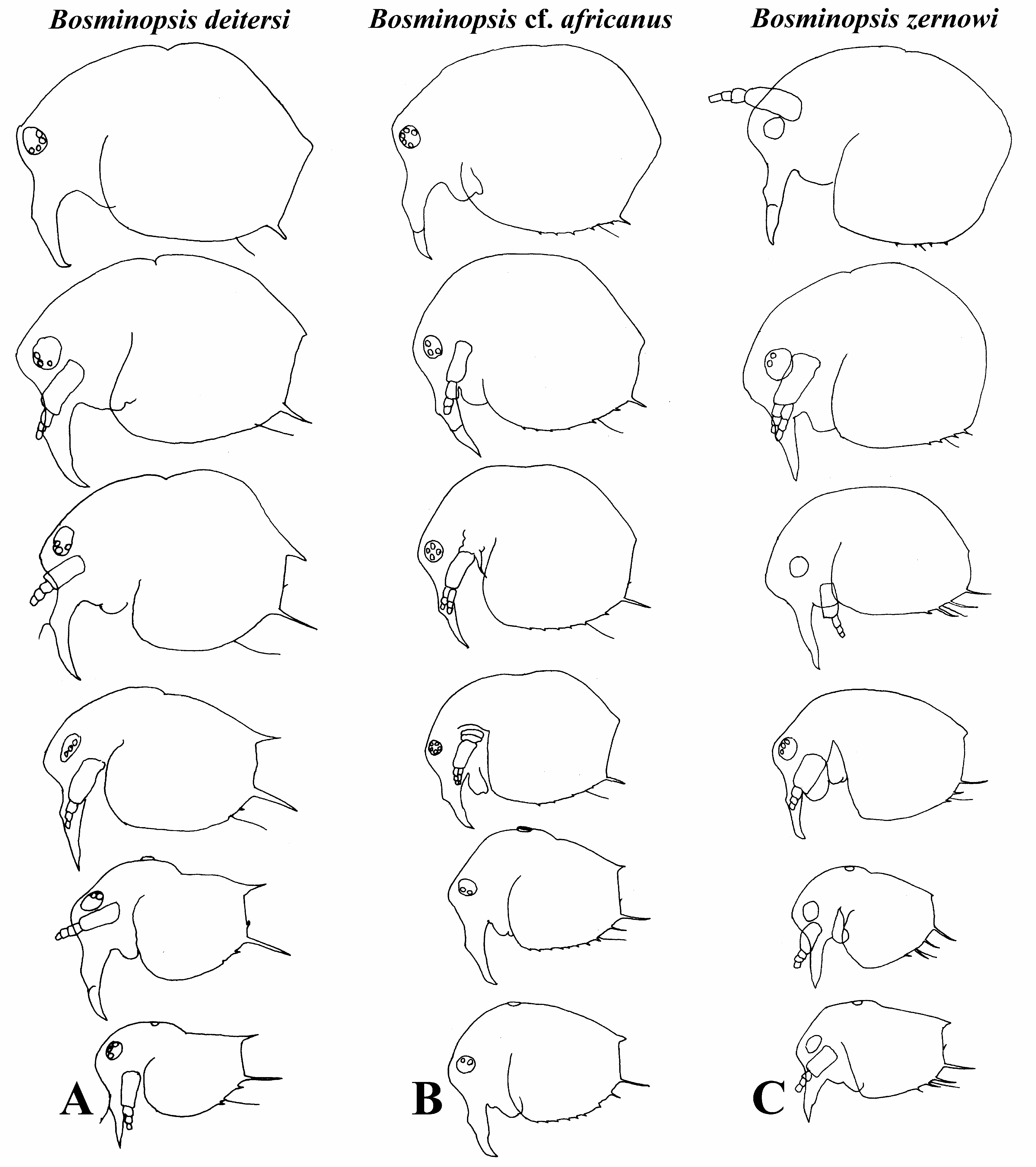
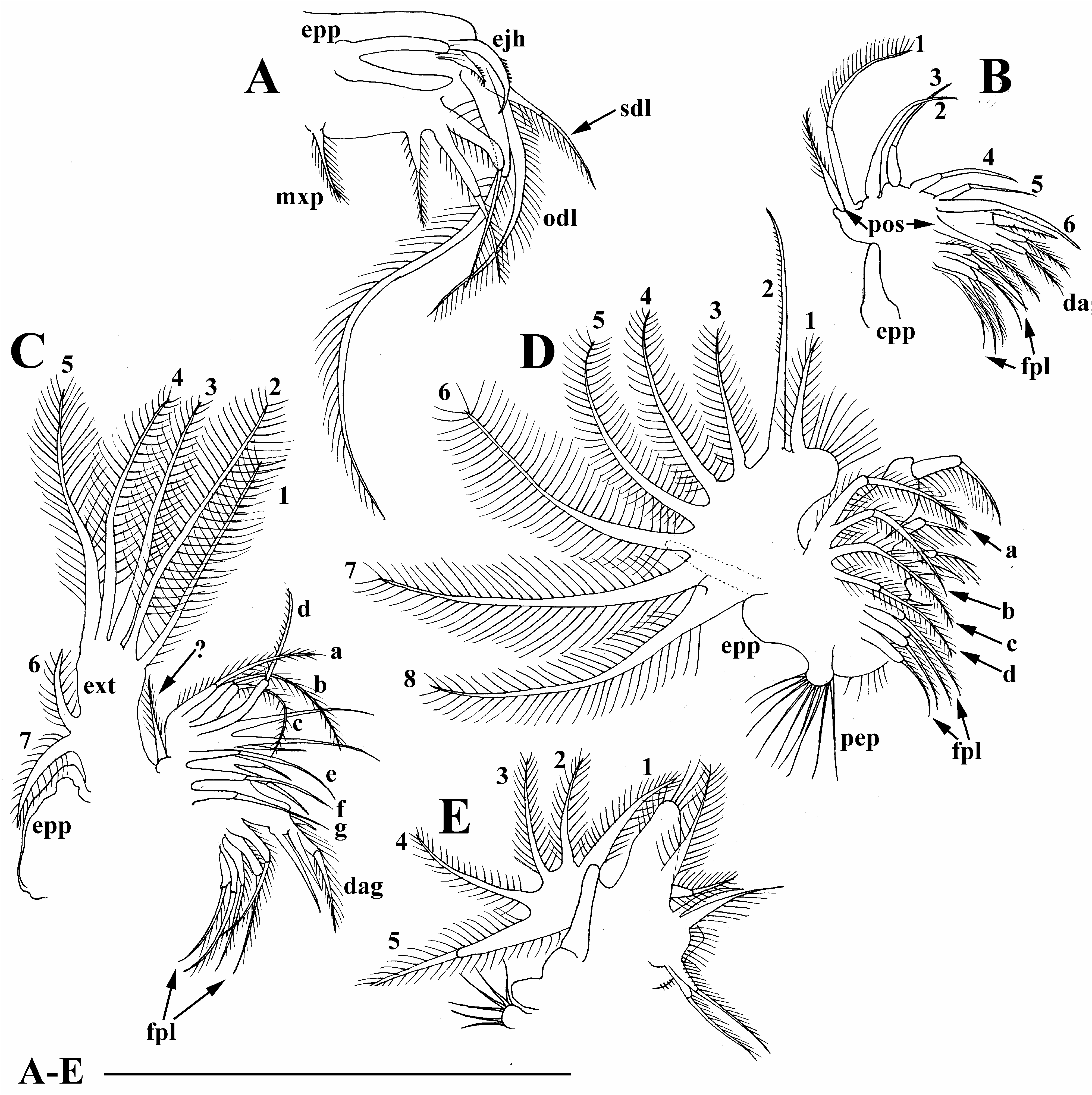
Diagnosis. Female. Body of parthenogenetic adult mostly subovoid ( Figs. 2A,B View FIGURE 2 ), juvenile female with lower posterior portion ( Fig. 2C View FIGURE 2 ). Postero-dorsal projection very short or absent. Reticulation in general not distinct, but well-expressed on head anterior face. Head with a small ocular dome; proximal parts of antennae I in females fused with each other and rostrum ( Fig. 3B View FIGURE 3 ). Exopod and endopod of antenna II with three segments ( Figs. 3C,D View FIGURE 3 ). Valves of adult with a single short mucro-like spine ( Figs. 2A,B View FIGURE 2 ), juvenile female with a single long mucro and a short mucro-like spine ( Figs. 3E–M View FIGURE 3 ). Maxilla I with three setae ( Fig. 4A View FIGURE 4 ). Five pairs of thoracic limbs ( Figs. 4B–I View FIGURE 4 ) as in other species of B. deitersi group, but inner-distal portion of limb III with six posterior (soft) setae ( Fig. 4E View FIGURE 4 : a–f). Postabdomen without an especial basis of postabdominal setae; basal spine on postabdominal claw very large (longer than half of claw length) ( Fig. 3N View FIGURE 3 ).
Ephippial female egg chamber sculptured by large polygons ( Figs. 2D,F View FIGURE 2 ), lateral keels not so sharp as in B. deitersi and not so large as in B. zernowi .
Adult male ( Fig. 2G View FIGURE 2 ) has a shallow depression between head and valves; postero-ventral angle with a single mucro; limb I with a copulatory hook relatively large and thick, and blunt tip.
Redescription based on the specimens from Thailand. Parthenogenetic female. Body in lateral view subovoid, depression between head and valves absent ( Figs. 2A,B View FIGURE 2 ). Dorsal margin regularly convex, postero-dorsal and postero-ventral angles rounded in old adults ( Fig. 2A View FIGURE 2 ), but expressed in smaller adults ( Fig. 2B View FIGURE 2 ) and juveniles ( Fig. 2C View FIGURE 2 ). Ventral margin from slightly convex to almost straight; anterior margin of valves slightly convex.
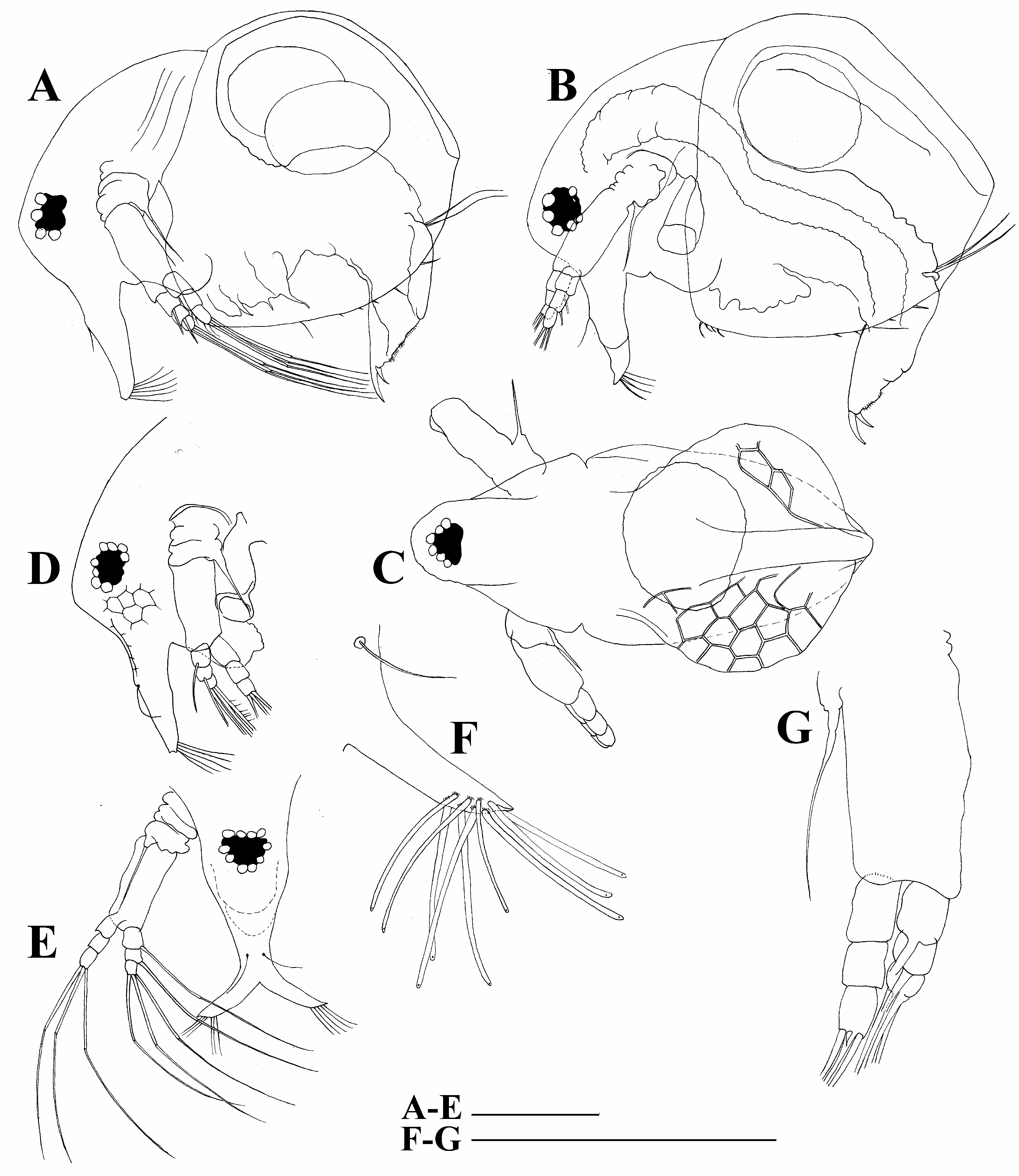
Head relatively large, with a low ocular dome, reticulation maximally expressed on its anterior face ( Fig. 2B View FIGURE 2 ). Compound eye of moderate size, ocellus absent. Lateral and dorsal head pores absent. Labrum with fleshy and round main body ( Fig. 2A View FIGURE 2 ).
Valve subovoid. Antero-ventral portion of valves with a series of setae ( Fig. 2A View FIGURE 2 ). Ventral margin slightly convex, with a series of small spinules and a long seta. Postero-ventral angle of valves with a single short mucro and sometimes with one additional relatively long spinule ( Figs. 3G–M View FIGURE 3 ).
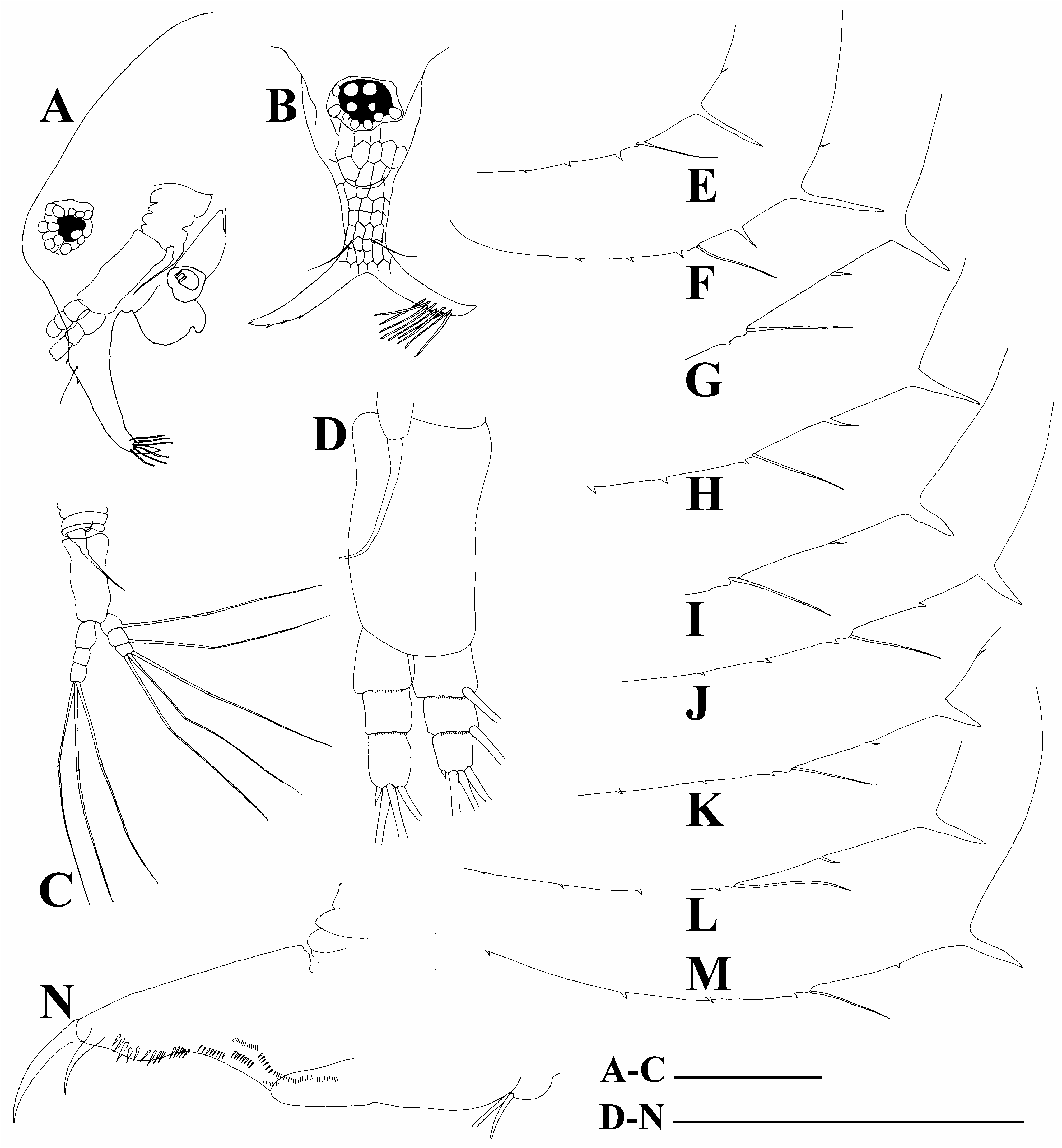
Antennae I in their proximal portions fused together and with rostrum ( Fig. 3A View FIGURE 3 ); lateral portions directed downwards and slightly curved ( Fig. 3B View FIGURE 3 ). Fused portion of antennae I with two antennular sensory setae. Distal portion of each antenna I with nine aesthetascs.
Antenna II coxal portion bearing a single long seta and a single minute seta on a conical elevation. Both exopod and endopod short, consisting of three segments. Antennal formula: setae 0-0-3/1-1-3; spines 0-0-1/0-0-1 ( Figs. 3C,D View FIGURE 3 ). All antenna II setae equal in size ( Fig. 3C View FIGURE 3 ).
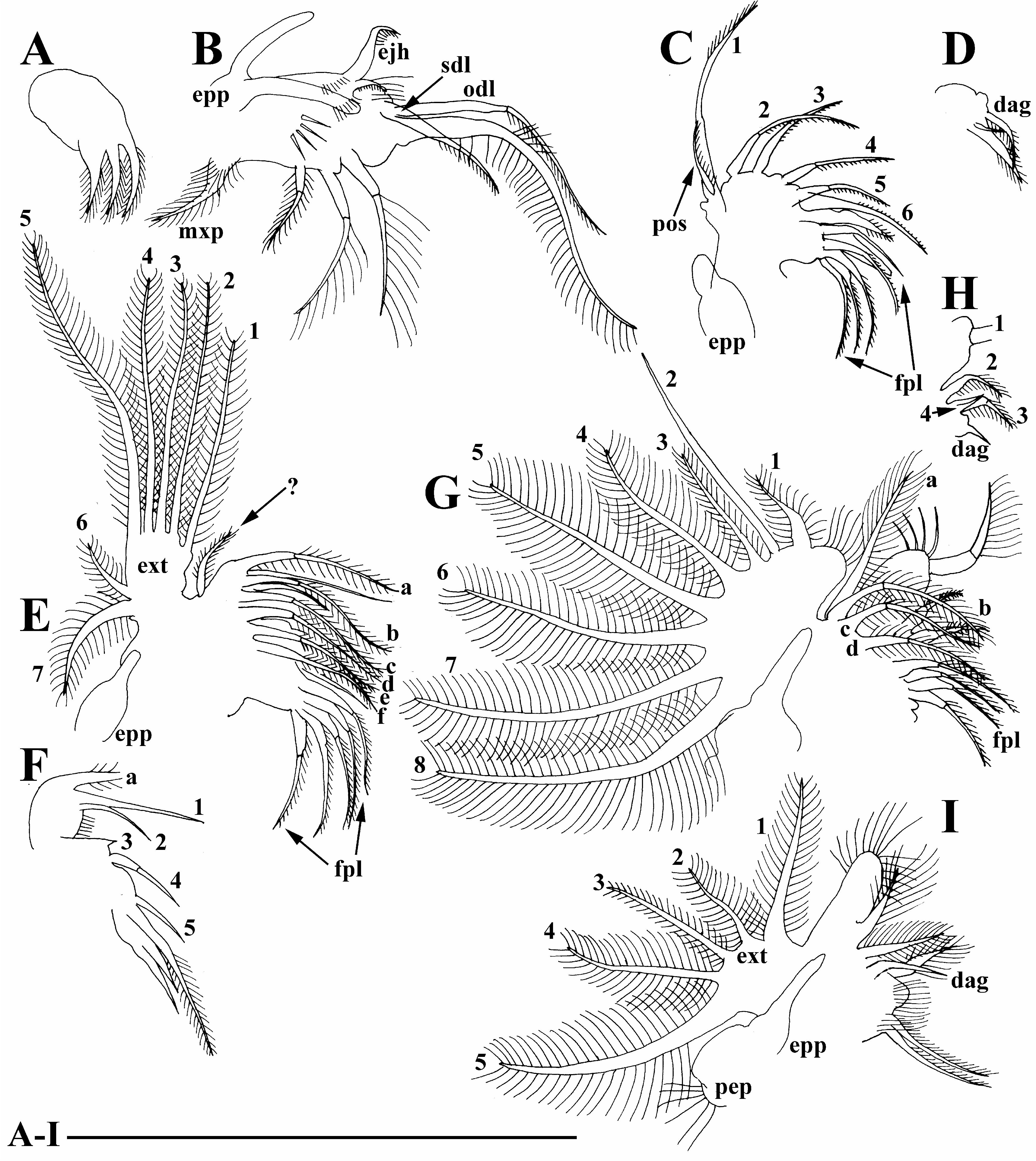
Maxilla as a small lobe with three setulated setae ( Fig. 4A View FIGURE 4 ).
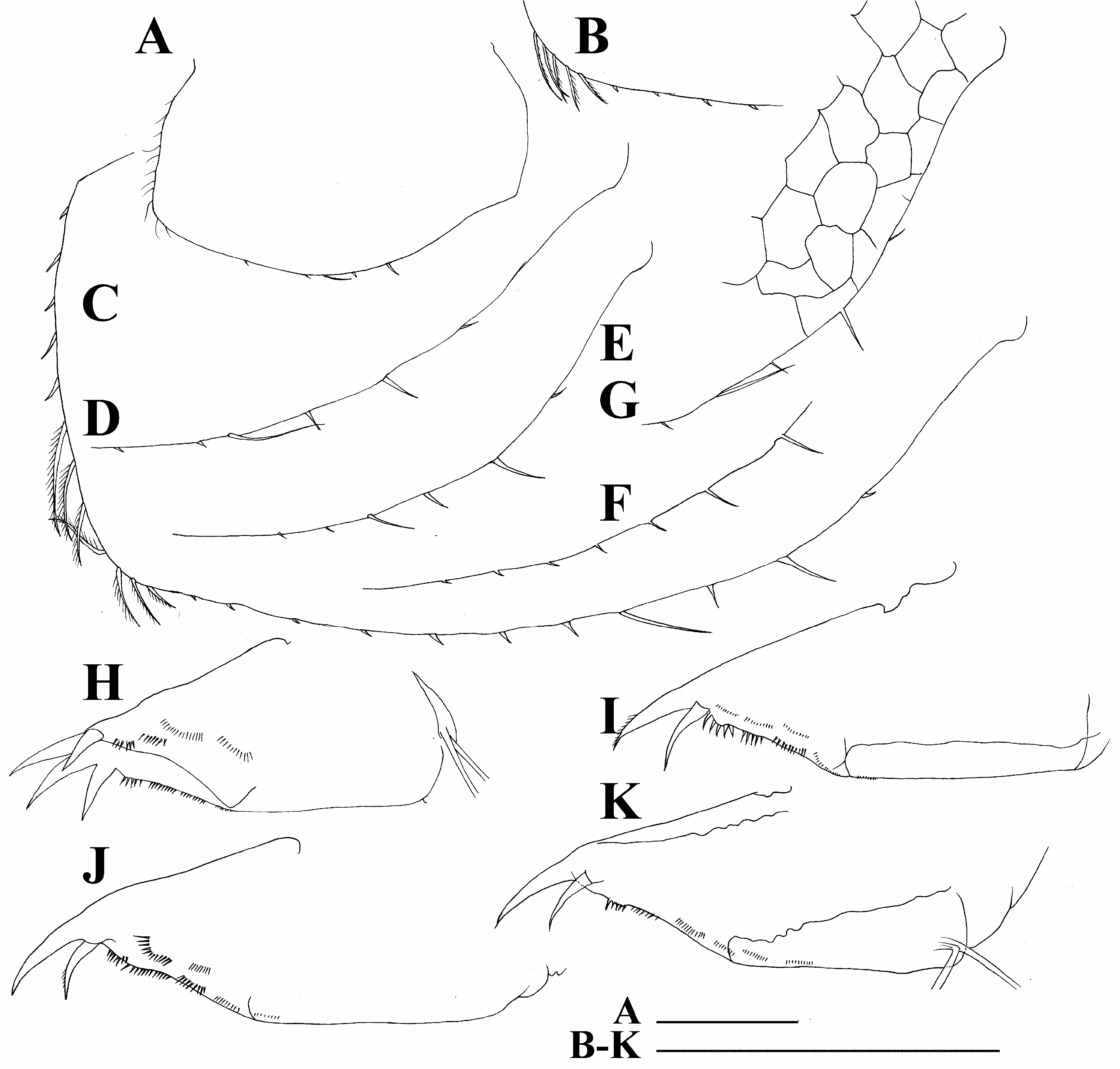
Limb I large, epipodite with two long finger-like projections ( Fig. 4B View FIGURE 4 , epp). Outer distal lobe ( Fig. 4B View FIGURE 4 , odl) with two long setae covered by setules. Inner subdistal lobe (in terms of Kotov, 1997a) bearing only a single setulated seta ( Fig. 4B View FIGURE 4 , sdl). Inner portion of limb corm with three soft setae, two equal in size and third one short; a bunch of long spinules on limb corm. Two ejector hooks different in size, robust, armed with small denticles ( Fig. 4B View FIGURE 4 , ejh). Maxillar process ( Fig. 4B View FIGURE 4 , mxp) bearing a single setulated seta.
Limb II small, epipodite with a single short finger-like projection ( Fig. 4C View FIGURE 4 , epp). Inner portion with six anterior setae (1–6) and two posterior setae ( Fig. 4C View FIGURE 4 , pos). Distal armature of gnatobase with three setae of different size ( Fig. 4D View FIGURE 4 , dag), filter plate with five setulated setae ( Fig. 4C View FIGURE 4 , fpl).
Limb III epipodite with a single finger-like projection ( Fig. 4E View FIGURE 4 , epp); exopodite rectangular ( Fig. 4E View FIGURE 4 , ext), bearing seven (two lateral and five distal) setae ( Fig. 4E, 1–7 View FIGURE 4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 5 View FIGURE 6 View FIGURE 7 ). Inner-distal portion with three anterior seta, seta 1 long, seta 2 shorter than seta 1 and seta 3 very small ( Fig. 4F View FIGURE 4 , 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ). Proximal endite with two anterior setae of same size ( Fig. 4F, 4–5 View FIGURE 4 View FIGURE 5 ). Posterior face of limb with six long soft setae ( Fig 4E, a–f View FIGURE 4 ) plus a seta of unclear homology (?). Distal armature of gnatobase with three setae ( Fig. 4F View FIGURE 4 ). Filter plate with five setae of similar size ( Fig. 4E View FIGURE 4 : fpl).
Limb IV epipodite supplied with a long finger-like projection. Exopodite semicircular, with eight long soft setae ( Fig. 4G, 1–8 View FIGURE 4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 ), seta 2 with very short setulation; inner distal portion with four anterior setae ( Fig. 4H, 1–4 View FIGURE 4 View FIGURE 1 View FIGURE 2 View FIGURE 3 ); distal most setae extra thick; posterior portion with four soft setae ( Fig. 4G, a–d View FIGURE 4 ). Distal armature of gnathobase reduced ( Fig. 4H View FIGURE 4 , dag), filter plate with four setae of similar size ( Fig. 4G View FIGURE 4 , fpl).
Limb V with a small, ovoid preepipodite ( Fig. 4I View FIGURE 4 , pep) and epipodite with a long finger-like projection ( Fig. 4I View FIGURE 4 , epp). Exopodite (ext) ovoid, with five soft setae (1–5) covered by long setules. Inner distal portion of limb as a densely setulated flat lobe bearing two soft setae. Distal armature of gnathobase ( Fig. 4I View FIGURE 4 , dag) with two setae, filter plate with two long setae.
Postabdomen compressed laterally, slightly narrow distally, without an especial inflated basis of postabdominal setae ( Fig. 3N View FIGURE 3 ), which are shorter than postabdomen ( Fig. 2A View FIGURE 2 ). Ventral margin straight. Preanal margin almost straight or slightly concave, without setules. Preanal angle expressed. Anal margin curved and postanal margin convex; both margins with series of a small spines. Postabdominal claw regularly curved, its basal spine long (about 2/3 of postabdominal claw length, or longer).
Juvenile female. Body with lower posterior portion, head relatively high. Dorsal head pore present only in instar I ( Fig. 6 View FIGURE 6 ). Postero-ventral margin of carapace with a single long mucro-like spine and a single long spinule ( Figs. 3E,F View FIGURE 3 ).
Ephippial female. Most of the body similar to parthenogenetic female, but dorsal portion of valves modified into ephippium ( Figs. 2D,F View FIGURE 2 ). Ephippium ovoid, sculptured with large polygons, with strong medial keel and paired lateral keels ( Figs. 2E,F View FIGURE 2 ). Ephippial chamber with a single egg.
Adult male. Unfortunately, only a single slide was available making impossible a detailed description. Body elongated, dorsal contour of head humped, dorsal contour of carapace almost straight, a slight depression between head and carapace. Head large, with specially large eye; ocular dam expressed; rostrum smooth ( Fig. 2G View FIGURE 2 ). Valve armed as in female; postabdomen similar to that of the female. Antenna I free in proximal portion; frontal seta long, located in middle of antenna I body. Antenna II with a long (as long as endopod), curved additional male seta on endopod apical segment. Limb with a well-developed copulatory hook.
Size. Parthenogenetic female 0.19–0.4 mm; ephippial female 0.30–0.32 mm; male 0.25 mm.
Variability. We have compared populations from Thailand with other populations from different regions. All studied populations from Laos, India and Vietnam have same features as populations from Thailand ( Fig. 5A–U View FIGURE 5 ). Juvenile stages with a single long mucro or mucro plus an additional relatively long spine on postero-ventral portion of valves; posterior portion of limb III with six soft setae.
Differential diagnosis. Populations of Bosminopsis from SE Asia belonged to Bosminopsis deitersi species group. We have concluded that all studied populations belong to a single species, Bosminopsis africanus , initially described from Africa ( Daday 1908), see below. Most part of North and East Eurasia is inhabited by another member of this group, B. zernowi . Original detailed drawings of this taxon are represented here ( Figs 6C View FIGURE 6 , 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 ), but we do not represent here its redescription as it was already given previously ( Garibian et al. 2021). Adequately described members of the B. deitersi group ( Kotov, 1997a; Garibian et al., 2021) are morphologically similar. Taxon from Oriental zone differs from B. zernowi in several features like a single long mucro-like (plus, sometimes, a relatively long additional spine) on postero-ventral portion of juvenile female valves; only one strong mucro-like spine in adult female; posterior portion of limb III with six soft, long setae (a–f); seta 1 of distal endite significantly longer than seta 2 (See Table 2 View TABLE 2 ). B. deitersi also have a single mucro-like spine on valves at adult and juvenile stages ( Fig. 6A View FIGURE 6 ). However, B. deitersi and B. zernowi have seven soft setae (a–g) on posterior portion of limb III and seta 1 have same size than seta 2 on distal endite of limb III. See Table 2 View TABLE 2 for some other characters.
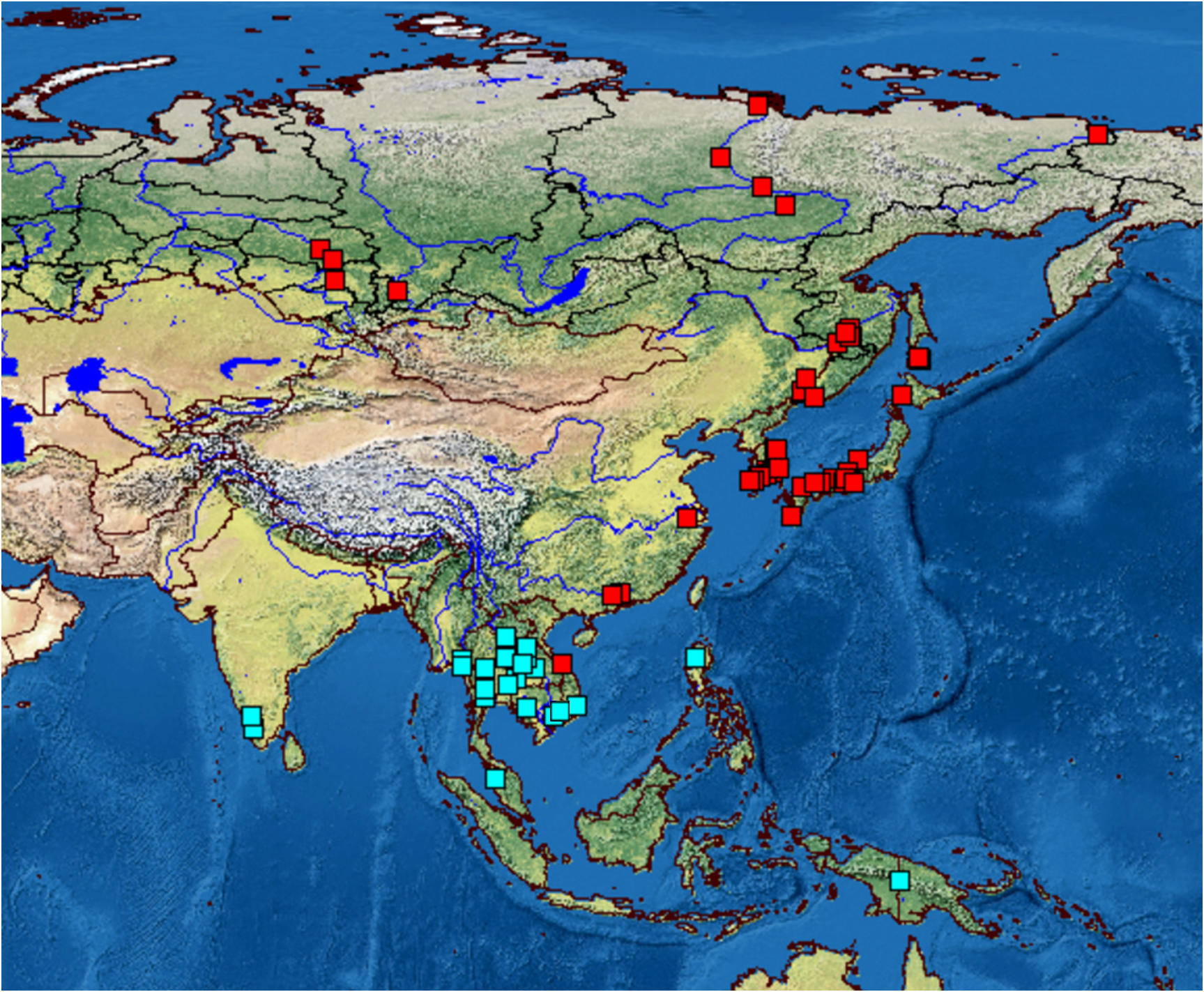
Distribution. B. africanus was recorded (as B. deitersi ) from many African countries (see Synonymy). We found this taxon in India, Myanmar, Cambodia, Thailand, Laos, Vietnam, Philippines, Malaysia and Papua New Guinea. Analysis of literature data (see Synonymy) confirms that is widely distributed through whole Oriental zone. It is present in the water bodies of different types, from large rivers, man-made reservoirs and large natural lakes to small ponds. But it is absent in temporary water bodies, probably resting eggs do not pass alive through drying.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |