
Globulidrilus helgei Christensen & Dózsa-Farkas, 2012
|
publication ID |
https://doi.org/10.11646/zootaxa.4000.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:DEB97452-A6E8-4AA9-A248-A65FD50D203F |
|
DOI |
https://doi.org/10.5281/zenodo.5202356 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC87B4-D128-FFF2-069E-93DEEC6BFD6F |
|
treatment provided by |
Plazi |
|
scientific name |
Globulidrilus helgei Christensen & Dózsa-Farkas, 2012 |
| status |
|
Globulidrilus helgei Christensen & Dózsa-Farkas, 2012 View in CoL
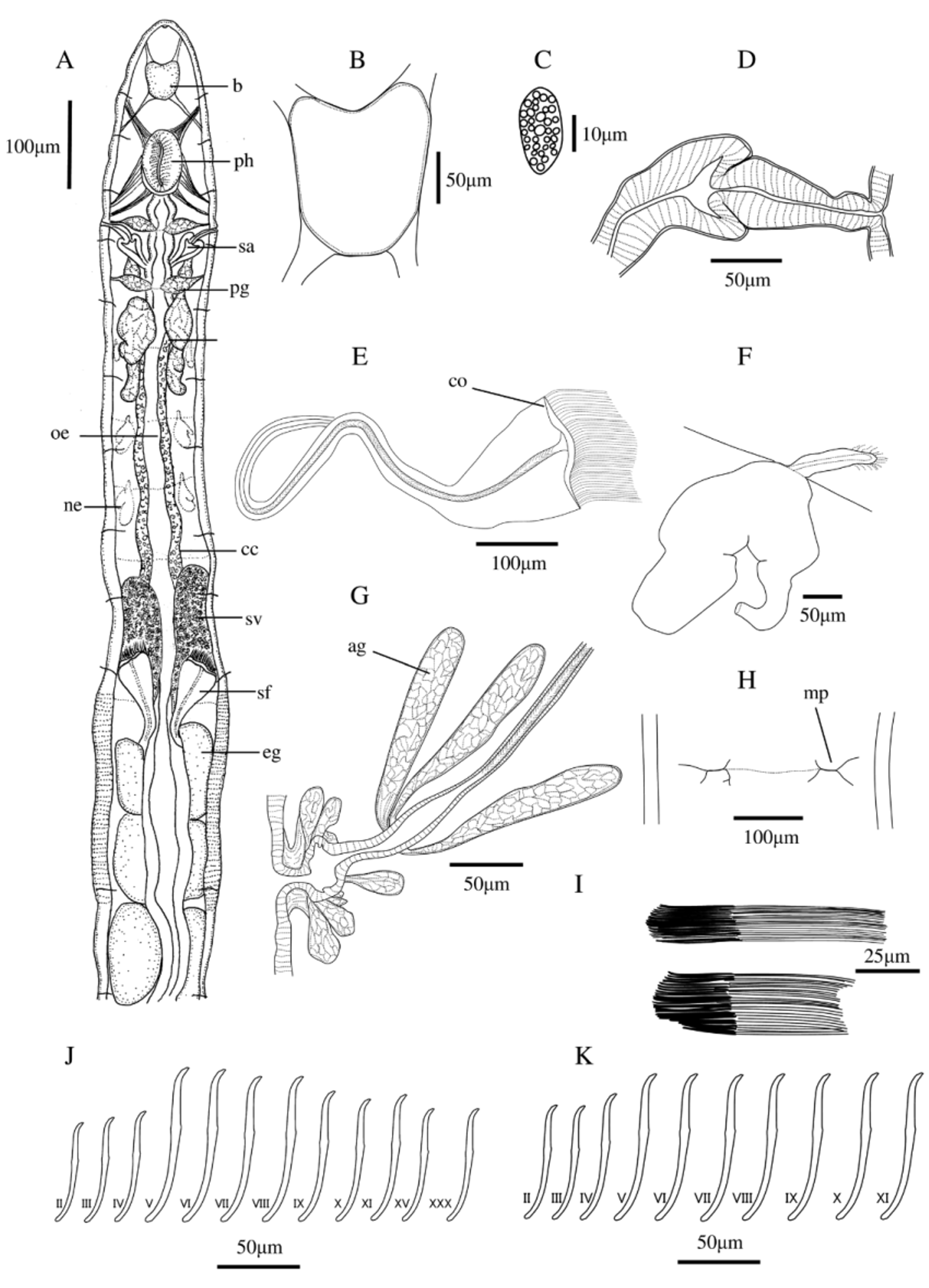
(Japanese name: kita-himemimizu, new) ( Figure 3 View FIGURE 3 )
Globulidrilus helgei Christensen & Dózsa-Farkas, 2012: 2770 View in CoL View Cited Treatment –2777 ( Figures 1 View FIGURE 1 –4: Table 2).
Material examined. Fourteen mature specimen, Mihon-en, Ozegahara mire, Katashina-mura, Tone-gun, Gunma Prefecture, Japan, N36°54’54.8’’, E139°11’53.7’’, 26 April 2014, T. Torii (NSMT-An 474–477). Three wholemounted mature specimens, same data as above, in the author’s collection.
Description of new material. Body whitish. Live length 5–8 mm, fixed length 4–7 mm. Width at clitellum 355 µm. Segment number 31–37. Chaetae slightly sigmoid, without nodulus, preclitellar chaetae 45.0–67.5 µm long, 4.0–5.0 µm wide, postclitellar chaetae 53.5–57.5µm long, 3.8–5.0 µm wide ( Fig. 3 View FIGURE 3 D,E). Chaetal formula 2–4 –2,3: 2–4–2,3. Length of preclitellar ventral chaetae varying in specimens, shortest mostly in II, maximum length mostly in IV–VIII. Postclitellar ventral chaetae almost as long as in preclitellar region. Clitellum over XII–1 /3XIII ( Fig. 3 View FIGURE 3 A), hyalocytes and granulocytes in dense rows, granulocytes dominate dorsally, the size of hyalocytes and granulocytes about the same, glands absent anteroventrally and between the penial bulbs but present posteroventrally.
Brain in I–II, dorsoventrally compressed and narrow, anterior part convex, posterior part deeply incised ( Fig. 3 View FIGURE 3 A), approximately twice as wide as anterior part, length 140–170 µm, posterior width 90–105 µm. Dorsal blood vessel originating in XIV or XV. Anterior blood system “lumbricilline” ( Coates & Ellis 1981), bifurcation in I ( Fig. 3 View FIGURE 3 A). Blood pale reddish. Paired postpharyngeal bulbs in III ( Fig. 3 View FIGURE 3 A). Transition of oesophagus to intestine gradual, no oesophageal appendages or intestinal diverticula. Pharyngeal glands three pairs, with ventral lobes, no secondary lobes, the third pair smaller than the others. Coelomocytes flattened, wavy outline with fine hyaline plasma threads, nucleate, vesicles blurred, ca. 2–3 times as long as wide ( Fig. 3 View FIGURE 3 C). Chloragocytes dense from VI, slightly smaller than coelomocytes. Preclitellar nephridia four or five pairs at 6/7 to 9/10 or 10/11, 130–145 µm long, sometimes the posterior pair absent or only on one side. Anteseptal part of nephridium large, including funnel plus canal, funnel comprising one-third or one-fourth of anteseptal part. Whole anteseptale about one-fourth or one-fifth the total length of nephridium. Postseptale without a distinct transition to efferent duct, large vesicle at the ectal opening. Postclitellar nephridia from 14/15.
Testes not observed. No seminal vesicle. Sperm funnel length 150–170 µm, width 55–65 µm, 3 times as long as wide. Collar not visible. Vas deferens narrowing from glandular part to male pore, moderately long, loose or tight, irregularly coiled in XII. Male glandular bulbs spherical. Thin bundles of filiform spermatozoa present in front of sperm funnel. Male glandular bulbs extension ca. one fourth or one fifth of body width, 80–90 µm long. Bursal slit 23–25 µm long. One spherical bulb-like structure in XII anteriorly, situated asymmetrically and resembling male glandular bulbs, 90–95 µm long, 80–85 µm wide. Oocytes in XI.
Paired spermathecal pores midlateral at 4/5, attached to oesophagus. Spermathecal ectal glands absent. Ampulla fusiform or oval ( Fig. 3 View FIGURE 3 A). Sperm as one or more ring-shaped bundles in ampulla lumen ( Fig. 3 View FIGURE 3 D).
Habitat. The specimens were present in the wet soil of the sphagnum mires.
Remarks. This is the first record of Globulidrilus helgei from Japan. The present material agrees with the original description of the species from Korea ( Christensen & Dózsa-Farkas 2012) 1994), except for (1) the lateral postclitellar chaetae which are slightly smaller than the anteclitellar ones (larger than anteclitellar chaetae in the original description of G. h e l g ei), (2) the presence of 4 or 5 pairs of preclitellar nephridia, from 6/7 to 9/10 or 10/11 (4 pairs, from 6/7 to 9/ 10 in G. h e l g e i), and (3) the length: width ratio of the sperm funnels, 3: 1 in the Japanese specimens and 4–6: 1 in the original description. Furthermore, in Korean G. h e l g ei the extra-bulbs in the male region were found in only half of the investigated specimens, and they were present in XI or in XI and XIII, whereas in the Japanese specimens they were present in ca. 70 % investigated specimens, and constantly in XII. These differences may indicate that the Korean and Japanese populations represent different species, but we prefer to collect more evidence (e.g. sequence data) before erecting a new species. Christensen & Dózsa-Farkas (2012) defined the genus Globulidrilus on the basis of characters of the chaetae, nephridium, spermatheca and others, and they transferred Marionina riparia Bretscher, 1899 , a freshwater and brackishwater species with worldwide distribution also found in Japan, to this genus, as Globulidrilus riparius .
Distribution. Korea ( Christensen & Dózsa-Farkas 2012), Japan (present study).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Globulidrilus helgei Christensen & Dózsa-Farkas, 2012
| Torii, Takaaki 2015 |