
HERPELIDAE, Laurent, 1984
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00838.x |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87B7-FFF9-FFB0-FC3B-95F5FDD365F5 |
|
treatment provided by |
Marcus |
|
scientific name |
HERPELIDAE |
| status |
|
HERPELIDAE View in CoL ( FIGS 4 View Figure 4 , S 7–S View Figure 7 8 View Figure 8 )
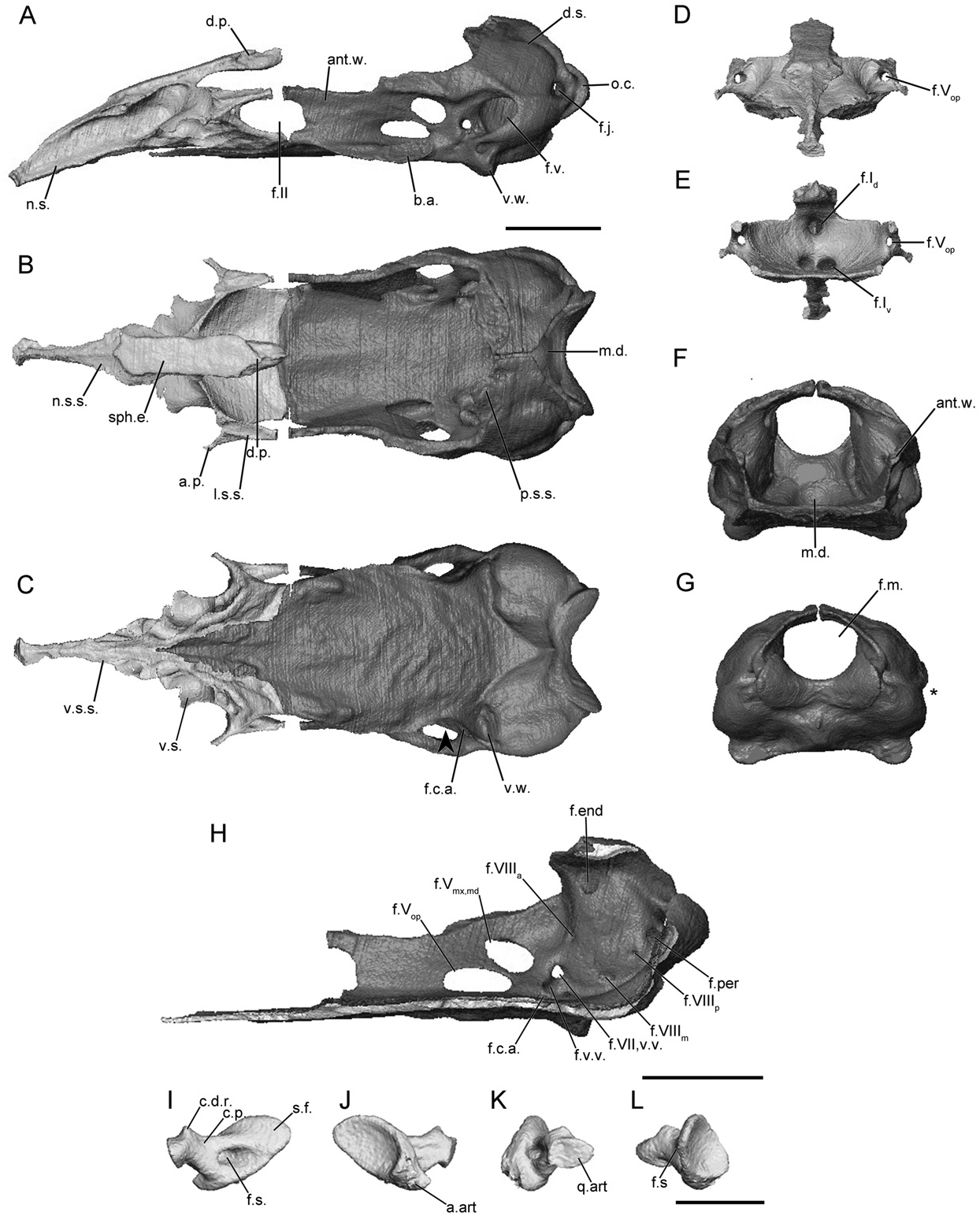
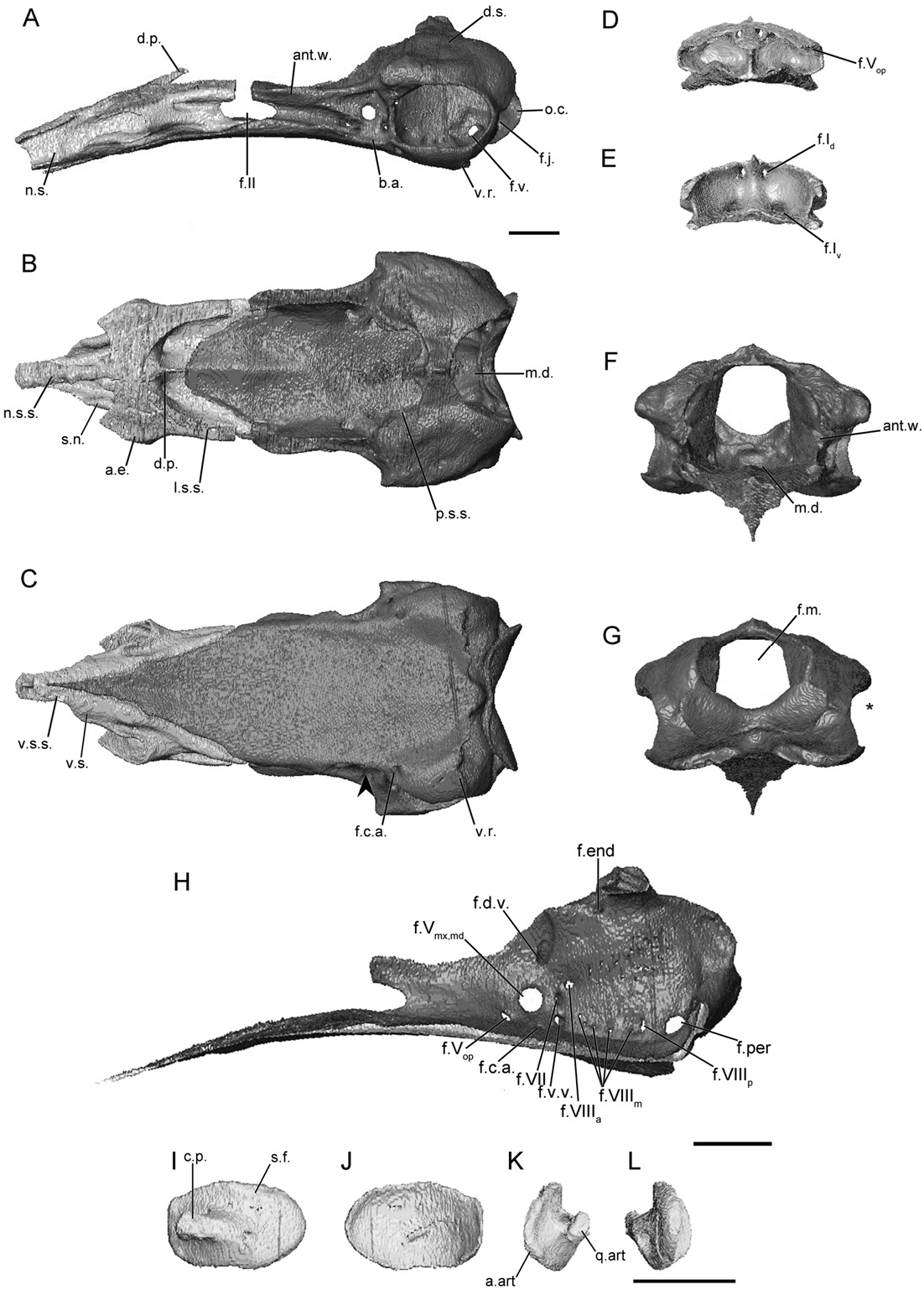
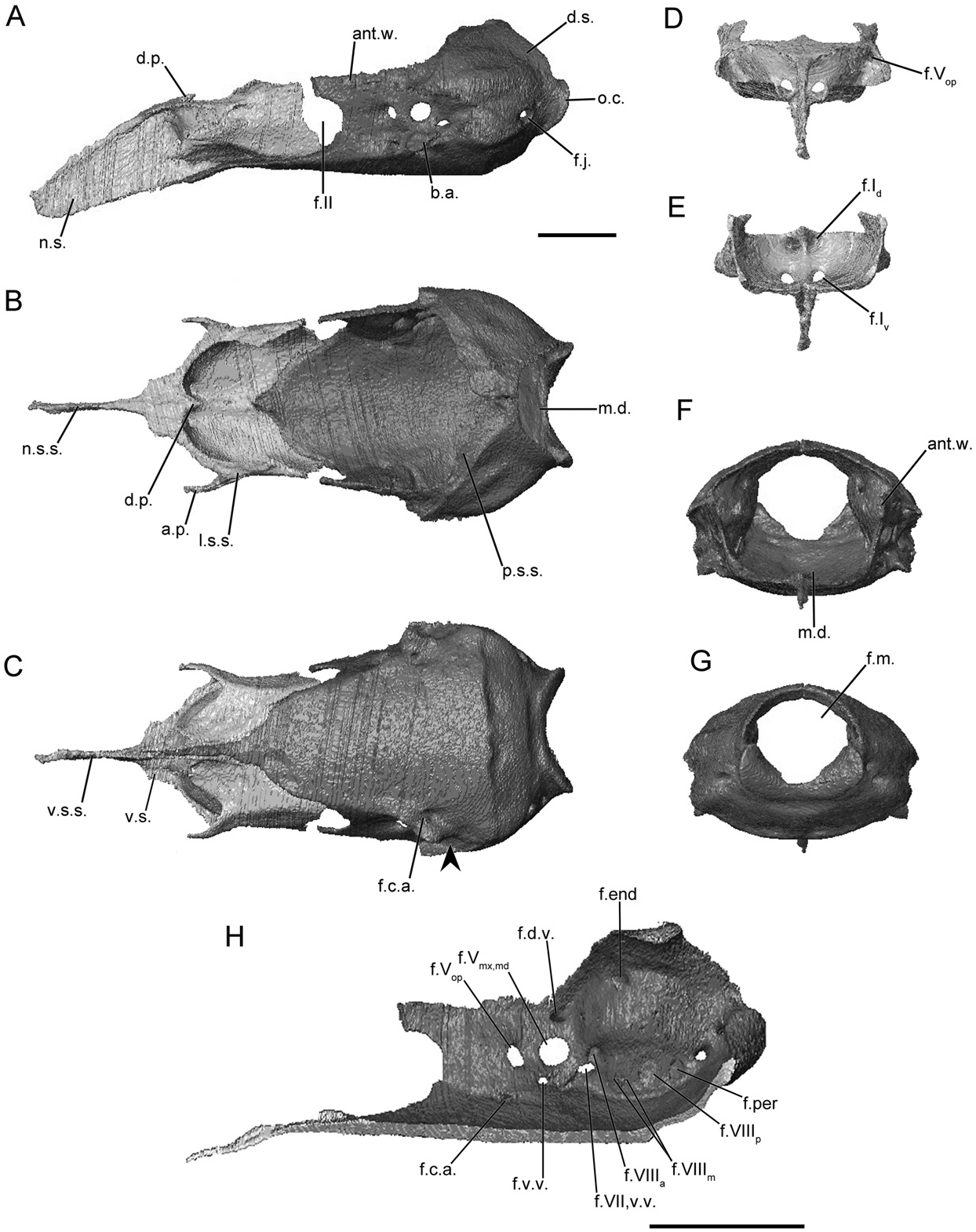
The main body of the sphenethmoid accounts for roughly 30% of the total length of the sphenethmoid in herpelids ( Fig. 4A View Figure 4 ). The lateral wall of the main body is short, and it is capped by a moderately broad sutural surface that receives an anterolateral lappet of the parietal ( Fig. 4B View Figure 4 ). An ossified thin, rod-like anterolateral process is present in herpelids, although it is variably developed in the specimens of He. squalostoma ( Fig. S8B View Figure 8 ). The dorsomedial process is broad. In Boulengerula boulengeri and He. squalostoma the process is exposed dorsally (as well as a region over the nasal septum) and reaches the level of the antotic wall of the os basale. It extends well beyond the antotic wall in He. squalostoma ( Fig. S8A View Figure 8 ). In lateral view the posterior margin of the lateral wall of the main body of the sphenethmoid is deeply incised by the optic foramen ( Fig. 4A View Figure 4 ). In He. squalostoma the outline of the foramen is anterodorsally oblique ( Fig. S8A View Figure 8 ). The posterior margin of the floor is moderately to gently concave in herpelids ( Fig. 4B View Figure 4 ).
The nasal region is divided by a long nasal septum ( Fig. 4A View Figure 4 ), which is dorsoventrally short, and tapers in height towards the tip of the snout ( Fig. 4A View Figure 4 ). In the species of Boulengerula it terminates in a small rounded tip ( Figs 4A, S View Figure 4 7A View Figure 7 ). The tip of the nasal septum closely approaches the anterior extent of the snout, with only a small gap separating it from the nasopremaxilla. The sutural surface over the nasal septum tapers to a thin, blade-like surface ( Fig. 4B View Figure 4 ). A thin shelf of bone extends laterally that forms a shelf that covers the ventral olfactory foramen dorsally and medially (weakly developed in He. squalostoma , along with a small shelf ventral to the olfactory foramen). This appears to differ from the solum nasi of other species.
The anterior wall contains paired dorsal and ventral foramina in He. squalostoma ( Fig. S8E View Figure 8 ), but only a single dorsal foramen and paired ventral foramina in the species of Boulengerula ( Figs 4E, S View Figure 4 7E View Figure 7 ). The dorsal foramen divides distally into the left and right foramina. The ventral pair of foramina is located close to either side of the midline, except for Boulengerula taitana in which they are more laterally placed ( Fig. S7E View Figure 7 ). The anterolateral foramen is present in the base of each anterolateral process ( Fig. 4D, E View Figure 4 ).
In dorsal view the antotic wall of the os basale of herpelids is bowed laterally at the level of the antotic foramina and is roughly parallel to the contralateral antotic wall anteriorly ( Fig. 4B View Figure 4 ). In anterior view the antotic wall of the os basale is orientated slightly dorsolaterally ( Fig. 4F View Figure 4 ). The dorsal sutural surface is slender for its entire length ( Fig. 4B View Figure 4 ). The anterior margin of the antotic wall is incised by the posterior margin of the oval optic foramen ( Fig. 4A View Figure 4 ), more deeply so in He. squalostoma ( Fig. S8A View Figure 8 ). The antotic wall contains three or four foramina ( Fig. 4H View Figure 4 ). This is referred to here as Pattern 5. The two trunks of the trigeminal nerve exit through their own large foramina. The facial nerve, along with a ventral vein, exit through an elongate foramen located posterior and ventral to the larger two ( Maddin, 2011). Variably an additional branch of the ventral vein is present and exits through a foramen slightly anterior to that for the facial and the other branch of the ventral vein in B. boulengeri ( Fig. 4H View Figure 4 ) and a dorsal vein and foramen is present in B. taitana ( Fig. S7H View Figure 7 ) and He. squalostoma ( Fig. S8H View Figure 8 ).
The dorsal surface of the otic-occipital complex is posteroventrally sloped. The dorsally exposed portion of this surface is broad in the species of Boulengerula ( Fig. 4B View Figure 4 ), but less so in He. squalostoma ( Fig. S8B View Figure 8 ). The fenestra vestibuli is small and has an ovoid outline. It is orientated anteroventrally and is located in the anterior-most portion of the otic capsule. In lateral view the occipital condyle protrudes well beyond the posterior limit of the otic capsules. A large jugular foramen is present, and is visible in lateral view ( Fig. 4A View Figure 4 ).
The medial wall of the otic capsule contains five to seven foramina. These include the four foramina common to all species examined here (endo- and perilymphatic foramina, anterior and posterior vestibulocochlear nerve foramina). Variably within the family one to three additional foramina are present that transmit the medial branch of the vestibulocochlear nerve ( Fig. 4H View Figure 4 ).
The anterior portion of the floor of the os basale is very narrow, and is triangular in outline ( Fig. 4C View Figure 4 ). The floor extends to reach the ventral surface of the nasal septum of the sphenethmoid ( Fig. 4C View Figure 4 ). A well-developed, medial depression is present in the posterior-most region of the floor of the os basale, just ventral to the foramen magnum ( Fig. 4B View Figure 4 ). Lateral to the antotic wall is a small, moderately well-developed basicranial articulation ( Fig. 4A View Figure 4 ). In ventral view the lateral margin of the floor of the os basale is moderately to weakly constricted medially, posterior to the basicranial articulation (arrowhead; Fig. 4C View Figure 4 ). A ventral projection is present on the ventral surface of the otic capsule. The posterior extent of the floor of the os basale terminates at a median point in B. boulengeri ( Fig. 4C View Figure 4 ) and a more rounded point in the other species. Only in B. taitana ( Fig. S7C View Figure 7 ) does the apex of the point approach the margin of the foramen magnum.
The foramen that leads to a canal for the carotid artery enters the ventral surface of the os basale ( Fig. 4C View Figure 4 ). This foramen is in a much more posterior and lateral location in B. boulengerula than in the other herpelids. Anteriorly the canal divides into lateral and medial canals, which in B. taitana and He. squalostoma terminate at foramina located ventrally to the trigeminal antotic foramina (and more posteriorly so in B. boulengerula ).
The footplate of the stapes is elongate anteroposteriorly ( Fig. 4I, J View Figure 4 ) and its long axis is inclined anteroventrally. The anterior margin of the footplate bears a large articular facet for contact with the margin of the fenestra vestibuli in the species of Boulengerula ( Fig. 4J View Figure 4 ). The medial surface of the footplate is strongly concave. The columellar process is long with a distal ridge ( Fig. 4I View Figure 4 ). A foramen pierces the base of the columellar process ( Fig. 4I View Figure 4 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |