
Fallomus ladakhensis Nanda & Sahni, 1998
|
publication ID |
https://doi.org/10.5281/zenodo.5377273 |
|
persistent identifier |
https://treatment.plazi.org/id/03DAFD42-FFA3-377B-FF7D-CCAAFC395C30 |
|
treatment provided by |
Marcus |
|
scientific name |
Fallomus ladakhensis Nanda & Sahni, 1998 |
| status |
|
Fallomus ladakhensis Nanda & Sahni, 1998
REFERRED MATERIAL. — 2 left mandibles with p4-m3 (SHM-CP 250, 353), 2 left m1s (TF 6181, 6182). The specimens are stored in the paleontological collections of the Department of Mineral Resources in Bangkok, Thailand.
LOCALITY. — Cha Prong pit of Nong Ya Plong coal mine, Phetchaburi Province, Thailand ( Fig. 1 View FIG ).
PALEOGEOGRAPHIC DISTRIBUTION. — Ladakh, northwestern Himalaya of India, Phetchaburi Province of Thailand.
AGE. — Late Oligocene.
EMENDED DIAGNOSIS. — Species slightly larger and significantly more hypsodont than Fallomus quraishyi Marivaux & Welcomme, 2003 . It differs from F. razae Flynn, Jacobs & Cheema, 1986 , F. ginsburgi Marivaux & Welcomme, 2003 and F. quraishyi in lacking the anterocingulid on molars, and in having more protruding labial and lingual cuspids that merge in generating a fully transverse bilophodont dental pattern. It differs from the previous species plus Diatomys Li, 1974 in showing the entoconid of both premolars and molars anteriorly displaced with respect to the hypoconid, cuspids less anteriorly inclined, and in having an enlarged hypoconulid twinned with the hypoconid, thus forming a small third posterior loph.
DESCRIPTION
Dental material
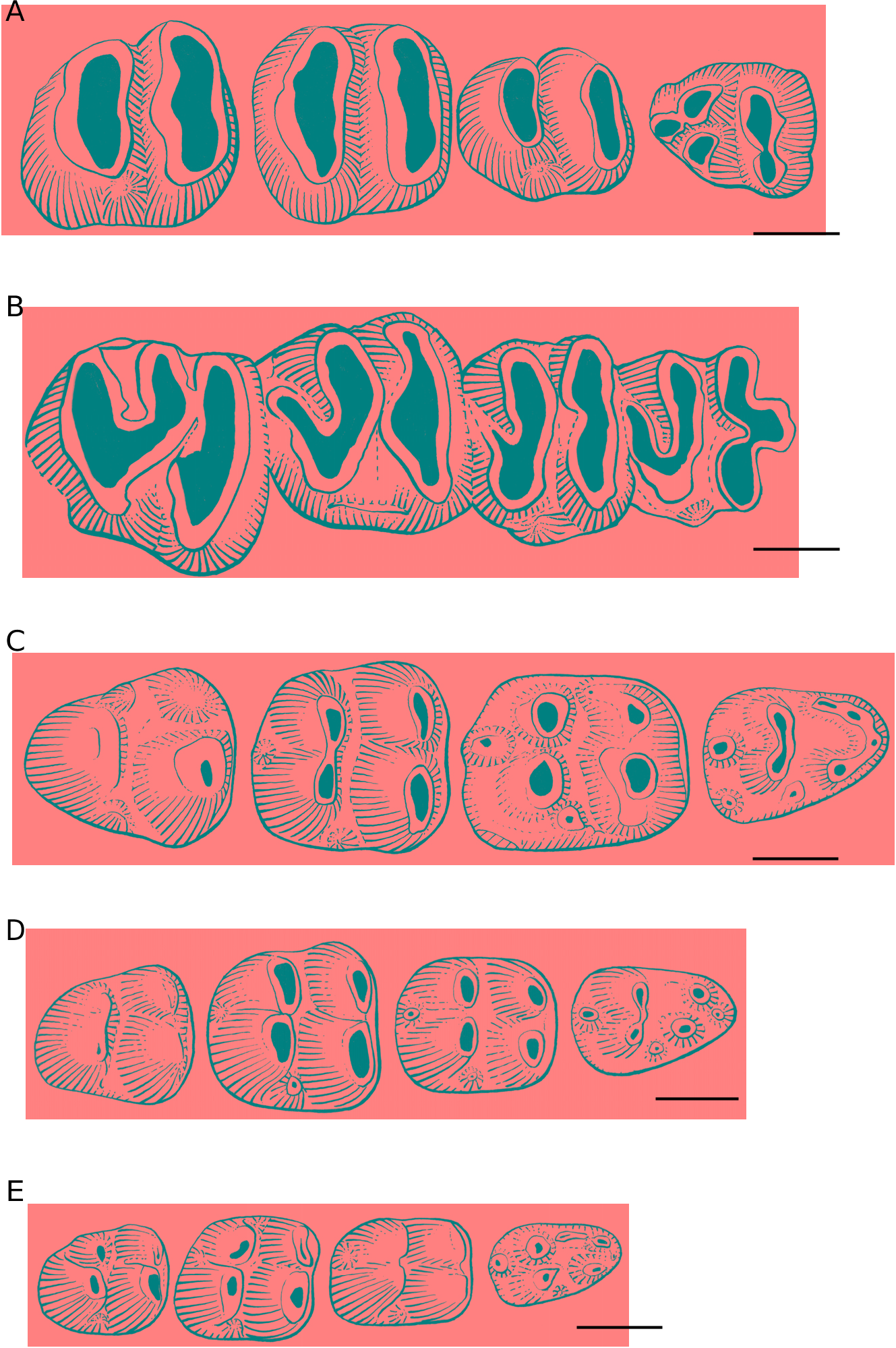
Lower premolar. The p4 is rectangular in outline and it is characterized by a strong anteroconid situated anteriorly between the protoconid and metaconid. This tooth is more labiolingually compressed on SHM-CP 250 ( Fig. 3C View FIG ) than on SHM-CP 353 ( Fig. 3A View FIG ). The hypoconid is the smallest cuspid of the tooth and connects both the inflated entoconid and the strong hypoconulid in a moderate stage of wear. The ectostylid is more salient on SHM-CP 353 than on SHM-CP 250.
First and second lower molars. m1 and m2 are similar in shape (oval-shaped) and in overall morphology, except that m2 is slightly larger ( Table 1) than m1, and shows an entoconid more anteriorly displaced with respect to the hypoconid. Both teeth have protruding labial (protoconid and hypoconid) and lingual (metaconid and entoconid) cuspids that merge labiolingually in generating a transverse bilophodont pattern from the first stages of wear. On the moderately worn and isolated specimen TF 6181 ( Fig. 3B View FIG ), the size of the cuspids can be better appreciated, thus showing that the hypoconid is smaller than the protoconid, metaconid, entoconid, and hypoconulid. There is no posterolophid linking the hypoconid to the hypoconulid. A “false” posterolophid appears artificially after wear by contact between the sides of the inflated hypoconid and hypoconulid, generating a small third posterior loph. On SHM-CP 353 ( Fig. 3C View FIG ), the hypoconulid of m1 and m2 is as large as the protoconid or the entoconid (as on the holotype WINF/A 1706 described by Nanda & Sahni [1998]; Fig. 4B View FIG ), whereas it is reduced on the m1 of SHM-CP 250 ( Fig. 3A View FIG ). The trigonid and the talonid do not show a significant difference of elevation, and they remain separated by the deeply incised transverse sinusid, which is labiolingually open due to the lack of an ectolophid. The protoconid and the metaconid are linked anteriorly by a metalophulid I that forms the anterior margin of the teeth. The protoconid does not show a development of its posterior arm (metalophulid II absent). The ectostylid, located anterolabially to the hypoconid, is nearly indistinct on the m2 of SHM-CP 250 ( Fig. 3A View FIG ), whereas it is very prominent on its m1, and on both teeth of SHM-CP 353 ( Fig. 3C View FIG ) .
Third lower molar. The m3 of SHM-CP 250 ( Fig. 3A View FIG ) is the largest tooth of the toothrow ( Table 1). This tooth is morphologically similar to m2, but differs in showing a larger hypoconulid that forms, in association with the hypoconid, a posterior lobe on the talonid (as on WINF/A 1707 described by Nanda & Sahni [1998]; Fig. 4A View FIG ). Compared with m2, the entoconid is more anteriorly located with respect to the hypoconid and it is nearly twinned with the metaconid but separated from it by a shallow groove corresponding to the labiolingually opened sinusid.
Incisor enamel microstructure
A fragment of incisor was removed from the mandible SHM-CP 353 ( Fig. 5A) and embedded in artificial resin. The fragment was subsequently polished longitudinally, then etched for 30 sec with H 3 PO 4 (37%) to make microstructural details visible. The specimen was examined under the scanning electron microscope at different resolutions ( Fig. 5 B-D).
In longitudinal section, the total enamel thickness is about 205 mm with 86% representing the portio interna (PI, inner region). The portio externa (PE, outer region), with radial enamel, is indeed very thin ( Fig. 5B). The PI consists of decussating layers of prisms appearing as Hunter- Schreger bands (HSB), 3-4 prisms wide per band ( Fig. 5C). The straight HSB are inclined at 50° to the enamel-dentine junction (EDJ). There is no well marked transition zone between adjacent decussating HSB. The interprismatic matrix (IPM) in the PI is thin and never surrounds the
B
prisms but appears as interrow sheets running at a high angle (> 80°, nearly rectangular) to the prism direction and anastomozing only rarely between the prisms ( Fig. 5C, D). Such a crystallite arrangement in PI determines typical multiserial enamel ( Korvenkontio 1934). The pattern of the IPM arrangement further typifies a derived multiserial condition designated as multiserial subtype III (see Martin 1994).
COMPARISONS
The dental morphology of both upper and lower teeth of the Oligo-Miocene Diatomyidae is primary characterized by the absence of transverse and longitudinal crests, the occurrence of an enterostyle and an ectostylid at the posterior lingual and labial margins of the internal sinus and external sinusid, respectively, and by the development of inflated cusps and cuspids that usually merge labiolingually in generating a transverse bilophodont occlusal pattern. Fallomus ladakhensis differs substantially from the other species of the genus ( F. razae ; F. ginsburgi ; F. quraishyi ; early Oligocene) and Diatomys (early middle Miocene; Li 1974; Mein & Ginsburg 1985, 1997) in showing a tendency to the acquisition of a trilophodont pattern on lower teeth as a result of the prominent development of the hypoconulid ( Figs 3 View FIG ; 4 View FIG ; 6B View FIG ). Although lacking in Diatomys ( Fig. 6A View FIG ), the hypoconulid occurs, however, in F. razae , F. ginsburgi and F. quraishyi , but it is moderately developed (as high as the ectostylid), or strongly reduced to nearly indistinct on some specimens ( Fig. 6 View FIG C-E). Such a strong and unequivocal development of the hypoconulid in F. ladakhensis is associated with the anterior displacement of the entoconid, notably on m3, which is very close to the metaconid ( Figs 3A View FIG ; 6B View FIG ). The entoconid is labiolingually opposed to the hypoconid in Diatomys and only slightly anteriorly displaced in the other species of Fallomus whatever the degree of development of the hypoconulid. F. ladakhensis differs significantly from the other early Oligocene species of the genus in having teeth clearly hypsodont ( Fig. 3D View FIG ). For instance, the ratio (R) of height to length of the moderately worn m1 (TF 6181) of F. ladakhensis is about 0.9, almost twice as hypsodont as F. razae , F. ginsburgi and F. quraishyi , which were actually relatively low crowned (R # 0.5). Diatomys liensis Mein & Ginsburg, 1985 appears also clearly high crowned with respect to the early Oligocene Fallomus forms, but compared with F. ladakhensis , it remains relatively less hypsodont (R # 0.7). F. ladakhensis also differs from the early Oligocene forms in the crystallite arrangement of the incisor enamel microstructure. In F. ladakhensis , the IPM organizes in interrow sheets running at a high angle (> 80°, nearly perpendicular) to the prism direction and anastomozes only rarely between the prisms (multiserial subtype III). Such an organization is substantially different from that observed for example in F. ginsburgi ( Fig. 7A, B View FIG ), where the IPM runs at an acute
A B C D
angle to the prism direction and anastomozes regularly between the prisms (multiserial subtype II). In terms of biomechanics, the crystallite arrangement typifying the subtype III is more efficient for strengthening the enamel in the third dimension. As a result, the subtype III is generally considered as the most derived state of multiserial HSB ( Martin 1994). In that respect, regarding the incisor enamel microstructure, F. ladakhensis is more advanced evolutionarily than F. ginsburgi . In contrast, the enamel microstructure of F. ladakhensis is very close to that observed in the incisors attributed to Diatomys liensis from the early middle Miocene ( Fig. 7C, D View FIG ), in having the same number of prisms per HSB, and in showing the same degree of complexity of the IPM (i.e. organization and orientation). F. ladakhensis lacks, however, the transition zones between HSB, which are clearly visible in Diatomys ( Fig. 7C View FIG ). In Diatomys , the prisms are more steeply inclined than the bands and they bend apically from one HSB to the next, thus generating transition zones.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.