
Fridericia cf. paroniana Issel, 1904
|
publication ID |
https://doi.org/10.11646/zootaxa.4496.1.27 |
|
publication LSID |
lsid:zoobank.org:pub:7C536E1E-5D5A-4E2D-9E4F-28F3CEA9664C |
|
DOI |
https://doi.org/10.5281/zenodo.5950217 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3D43A-E457-FFB1-2580-FA04FB40F844 |
|
treatment provided by |
Plazi |
|
scientific name |
Fridericia cf. paroniana Issel, 1904 |
| status |
|
Fridericia cf. paroniana Issel, 1904 View in CoL
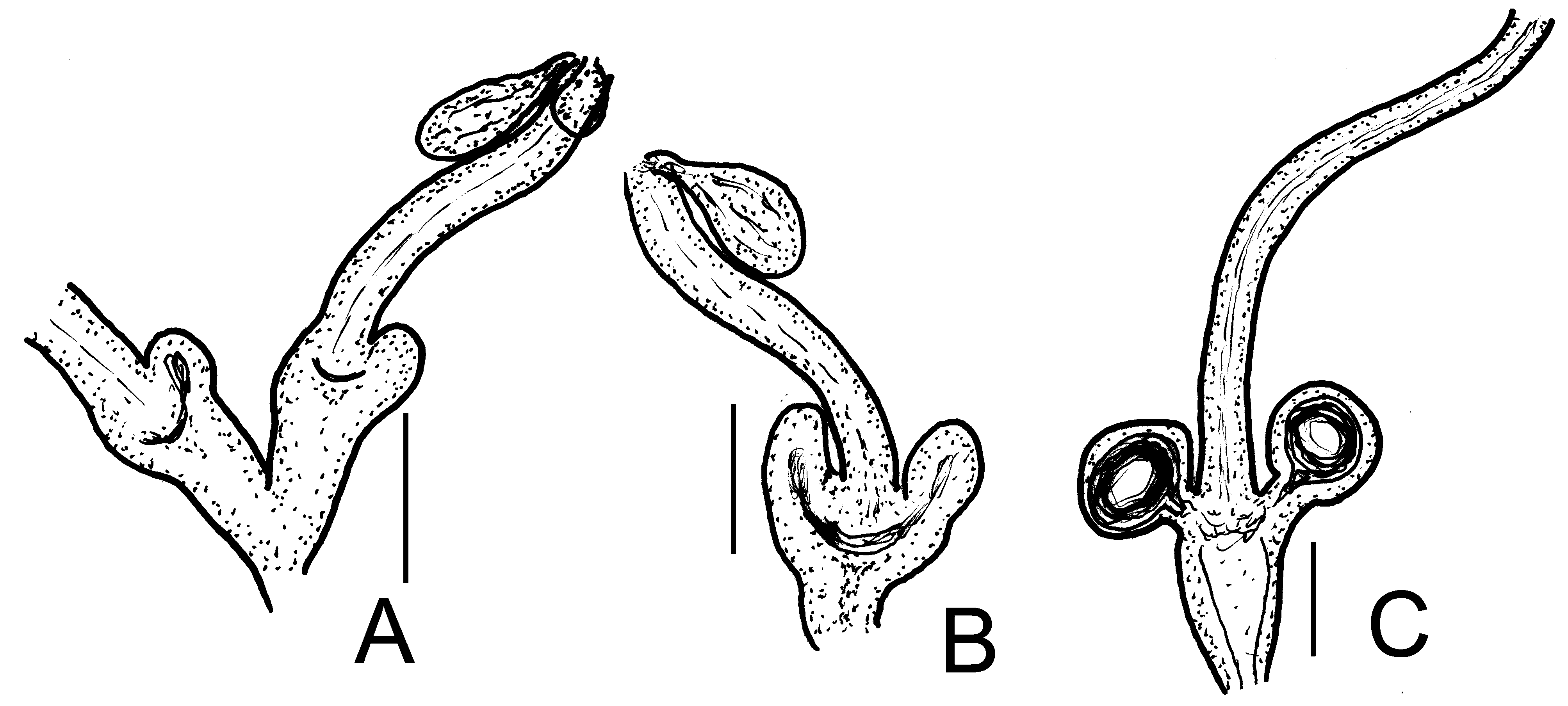
( Figures 19A,B View FIGURE 19 , 20 View FIGURE 20 )
Material examined. 14 specimens (two were deposited in the National Institute of Biological Resources , Korea NIBRIV0000810600, slide No. 2369, NIBRIV0000810601, slide No. 2378, NIBRIV0000810602, slide No. 2379).
Description. Middle large worms, body length 8.5–12.5 mm, width 240–330 µm at VIII and 300–350 µm at the clitellum in vivo. Length of fixed specimens 7.0– 9.5 mm, width 210–330 µm at VIII and 23 0–360 µm at the clitellum. Segments 39–47. Chaetal formula: 2 – 2: 2 – 2, in the segment XII absent. Chaetae in bundles: 38–45 × 5 µm preclitellarly, at the body-end 40– 55 x 5 µm. Head pore at 0/I. Dorsal pores from VII. One or two large epidermal glands in transverse rows ( Fig. 19C View FIGURE 19 ). Clitellum in XII–1 /2XIII, girdle-shaped, hyalocytes and granulocytes arranged in dense or indefinite rows, between bursal slits only granulocytes ( Fig. 20A–B View FIGURE 20 ). Body wall 17–25 µm, cuticle variable, 1–2.5 µm, fixed.
Brain egg-shaped, about 170 µm long in vivo (115 µm, fixed) 1.5–2 times longer than wide. Oesophageal appendages a-type, with wide lumen mostly without branches ( Fig. 20E View FIGURE 20 ). Pharyngeal glands ( Fig. 20F View FIGURE 20 ) in IV mostly with dorsal connection and without ventral lobes. Pairs in V with or without dorsal connection with ventral lobes, pair in VI free in dorsally, with large ventral lobes. Chloragocytes from V, 20–23 µm long, fixed. Dorsal vessel from XVIII–XX, blood colourless. Midgut pars tumida between XXVII–XXXVI, in 4–5 segments. Five pairs of preclitellar nephridia from 6/7 to 10/11, length ratio anteseptale: postseptale about 1:2, midventral origin of efferent duct. Coelomo-mucocytes numerous, mucocytes type a (length 21–34 µm in vivo, 25–33 µm, fixed), lenticytes large, 9–13 µm long ( Fig. 20D View FIGURE 20 ). Chylus cells between XII–XVI, occupying 2–3 segments ( Fig. 20H View FIGURE 20 ). Seminal vesicle absent. Sperm funnels ( Fig. 20G View FIGURE 20 ) cylindrical, approximately 90–105 µm long ( in vivo and fixed), about twice as long as wide, collar slightly narrower than funnel body, spermatozoa about 115 µm long, heads 37– 40 µm long, fixed. Diameter of sperm ducts 6–8 µm, fixed. Male copulatory organs 70–90 µm long, 45–70 µm wide and 40–60 µm high, fixed ( Fig. 20B View FIGURE 20 ). Bursal slits T-shaped ( Fig. 20I View FIGURE 20 ). Subneural glands absent. Spermathecae ( Figs. 19A–B View FIGURE 19 , 20K–L View FIGURE 20 ) variable: sometimes with "typical" shape: ectal ducts about 105–190 µm long and 17–25 µm wide in vivo (80–120 µm long, 14–18 µm wide, fixed), with large, floppy ectal gland (40–62 µm long, fixed) ( Fig. 20L View FIGURE 20 ); other specimens with another smaller gland (25–30 µm) ( Fig. 20J View FIGURE 20 ). Ampullae about 30–38 µm wide (fixed), with two small diverticula and separate openings into oesophagus dorso-laterally ( Fig. 20K View FIGURE 20 ). Other specimens with only one diverticulum of each ampulla or diverticula very small, even virtually not observable. In some specimens, the proximal parts of spermathecae fused entally, with a common opening into oesophagus dorsally ( Fig. 20L View FIGURE 20 ). One to three mature eggs at a time.
Distribution and habitat. In Mt. Hallasan, only one locality: site 14, in soil and litter layers of Sasa quelpaertensis forest.
Remarks. Despite the differences, we think that these specimens studied now from Korea can be considered as Fridericia paroniana , but due to differences we mark it as “ Fridericia cf. paroniana ”. The size, the segment number are about the same (the body diameter slightly thicker), chylus cells similarly in XII–XVI (2–3 segments long), midgut pars tumida in our specimens in XXVII–XXXVI (4–5 segments long), in F. paroniana in XXIII– XXXIV (4–5 segments long) (observation of Dózsa-Farkas on Hungarian specimens). Different traits are: dorsal vessel origin in XVIII–XX (XV–XVI in F. paroniana by Schmelz 2003, but in some Hungarian specimens also were observed in XIX by Dózsa-Farkas). We detected in the F. cf. paroniana more (1–3) mature eggs at a time, but only one egg by Schmelz (2003). The main differences are the variations of spermathecae, but individuals with only one diverticulum often occur also in the European populations, as given in the original description. However, a joint opening of the spermathecae into the oesophagus has not yet been observed in F. paroniana . For the time being we maintain that, based on other characteristic traits (abundant a-type mucocytes with large lenticytes, girdle-shaped clitellum, where between the bursal slits are only granulocytes, two chaetae in all chaetal bundles, the same form of pharyngeal glands and oesophageal appendages) the Korean specimens belong to Fridericia paroniana . We aim to solve the problem with a molecular comparative study in the near future.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |