
Achaeta koreana, Dózsa-Farkas & Felföldi & Nagy & Hong, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4496.1.27 |
|
publication LSID |
lsid:zoobank.org:pub:7C536E1E-5D5A-4E2D-9E4F-28F3CEA9664C |
|
DOI |
https://doi.org/10.5281/zenodo.5950199 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3D43A-E44B-FFA8-2580-F9B3FEE2F9AC |
|
treatment provided by |
Plazi |
|
scientific name |
Achaeta koreana |
| status |
sp. nov. |
Achaeta koreana View in CoL sp. n.
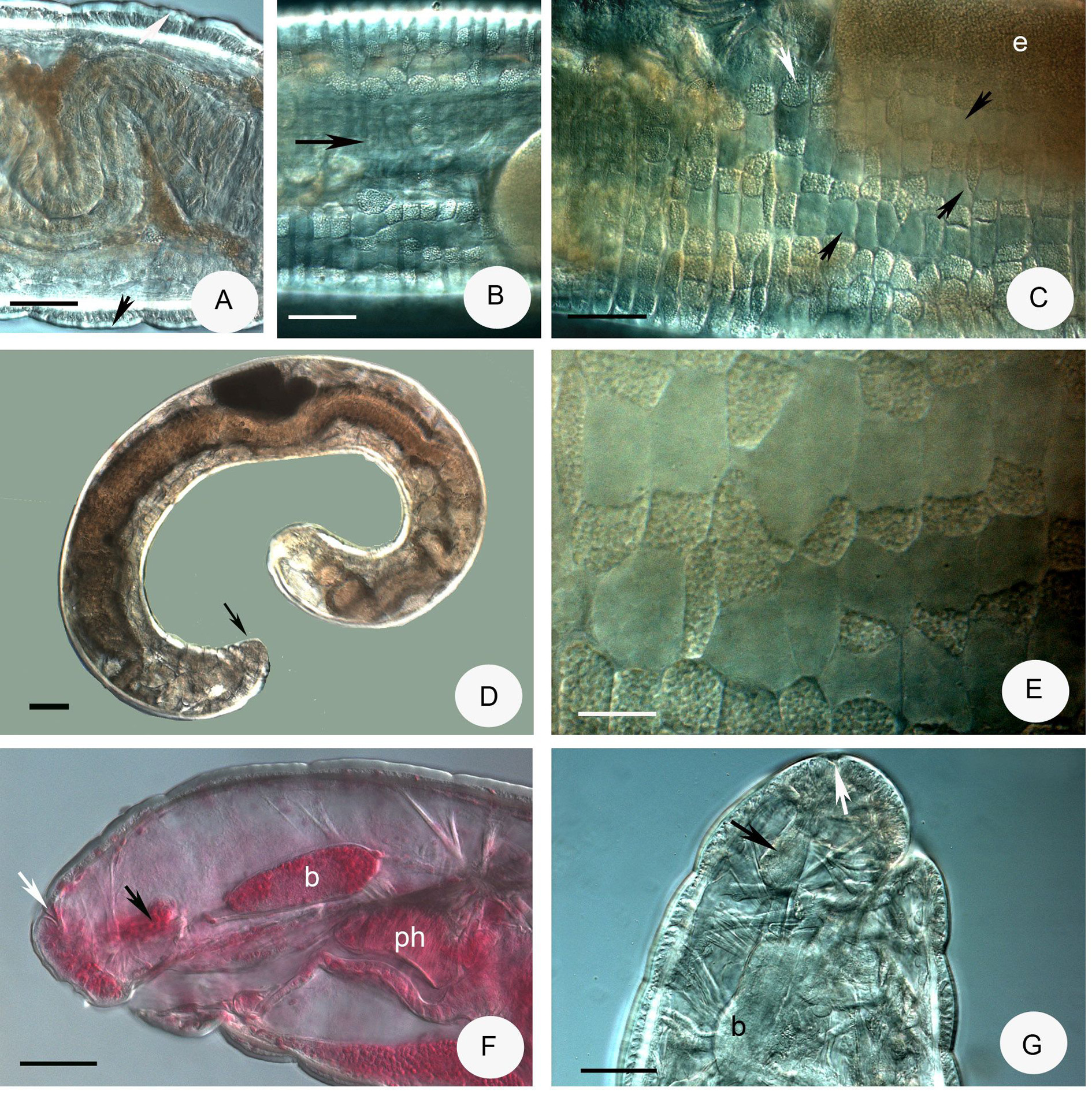
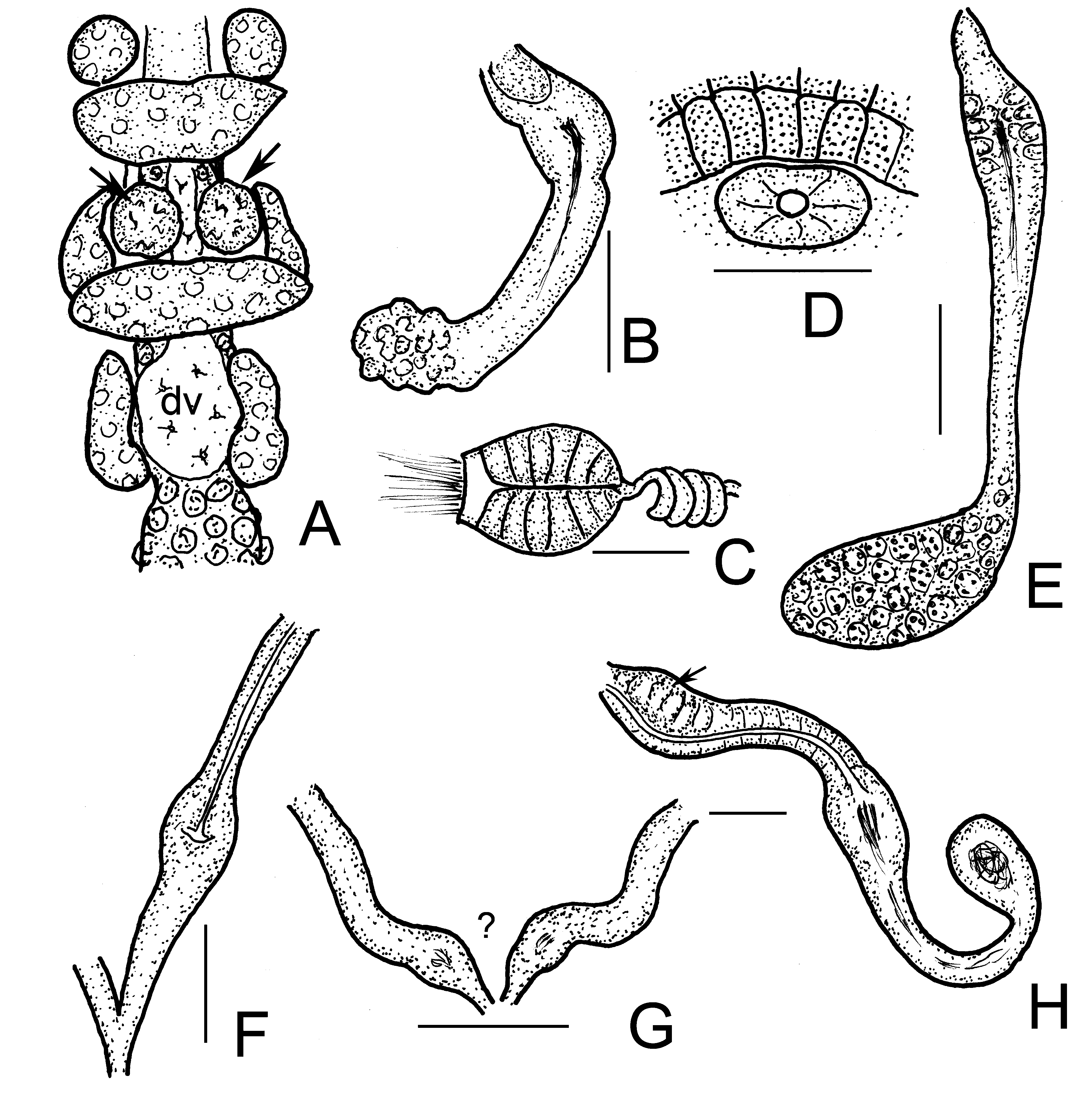
( Figures 1 View FIGURE 1 , 2 View FIGURE 2 , 5A–C View FIGURE 5 )
Type material. Holotype: NIBRIV0000810584 slide No. 2210, adult, stained whole mounted specimen. Type locality: site 4, Baekrokdam crater on the summit of Mt. Hallasan, Jeju Island, Korea, soil of grass on highland in North slope (N 33˚21'46.0", E 126˚31'58.0"), 1843 m elevation, 0 9.06.2016, leg. Y. Hong. Paratypes (in total 9 stained, adult specimens on slides and 37 specimens in 70% ethanol): NIBRIV0000810585, slide No. 2198, site 1, NIBRIV0000811380, slide No. 2276, site 2, P.113.1. slide No. 2212, from type locality, P.113.2.1–113.2.3. slides No . 2193–2195, site 6, P.113.3. slide No. 2196, site 5, P.113.4.1, slides No. 2197, site 1. In 70% ethanol: P.113.5, from type locality 10 specimens; P.113.6 site 2 seven specimens; P.113.7 site 5 eight specimens; P.113.8, site 6 twelve specimens.
Further material examined. 15 specimens investigated in vivo, 3 of them processed for DNA analysis.
Etymology. Named after the country where it was found.
Diagnosis. The new species can be recognized by the following combination of characters: (1) small, stout worms ( 2–3 mm long and 200–280 µm wide at clitellum in vivo), segments 17–22; (2) no pyriform glands; (3) clitellum developed only laterally: dorso-lateral area with hyalocytes in disordered three or four longitudinal rows; (4) dorsal blood vessel from VI; (5) pharyngeal glands at 4/5–5/6 connected dorsally, at 6/7 separate dorsally, the first ventral lobes separate from the dorsal lobes in IV, secondary ventral glands absent; (6) three pairs of preclitellar nephridia; (7) pars tumida of midgut from XIII–XIV, extending over 2 segments; (8) sperm funnel small, barrel-shaped, collar narrower than funnel body; (9) male pores in XII, ventrolaterally, no glandular body, only the pore surrounded by small inconspicuous glands; (10) spermathecae confined to V.
Description. Small worm ( Fig. 1D View FIGURE 1 ). Holotype 2.8 mm long, 275 µm wide at VIII and 315 µm at clitellum (fixed), 21 segments. Length of paratypes 2.5–3.8 mm, width 200–250 µm at VIII and 220–280 µm at clitellum in vivo, length of fixed specimens 2–3 mm, width 220–270 µm at VIII and 2 30–300 µm at clitellum, segments 17–22. No pyriform glands. Body wall thick, thickness dorsally 22–24 µm, cuticle 4–7 µm, ventrally 17–18 µm and 2–3 µm, respectively, in vivo ( Fig. 1A View FIGURE 1 ). Clitellum in XII–1 /2 XIII developed only laterally, dorso-lateral areas with hyalocytes in two or three disordered longitudinal rows ( Fig. 1C View FIGURE 1 ), hyalocytes large and conspicuous, 28–32 µm high and 17–19 µm wide, granulocytes 20–24 by 16–14 µm ( Fig. 1E View FIGURE 1 ). Ventro-laterally only granulocytes. The dorsal interruption 60–70 µm wide in vivo, dorsal borderline consisting of granular cells only in one longitudinal row ( Fig. 1B View FIGURE 1 ). Ventral borderlines inconspicuous. Head pore on the top of prostomium ( Figs. 1F–G View FIGURE 1 ). Spermathecal pores at 4/ 5 in lateral position. Male pores in XII.
Brain egg-shaped 1.5 times longer than wide, 80–90 µm long, fixed ( Fig. 1F View FIGURE 1 ). The prostomial ganglion ( Figs. 1F,G View FIGURE 1 ) well visible, 37–40 µm long (fixed). Pharyngeal glands at 4/5–6/7 ( Figs. 2A View FIGURE 2 , 5A View FIGURE 5 ), all with ventral lobes. The first and secondary dorsal lobes united dorsally, the third pair separate dorsally. Three pairs of preclitellar nephridia at 6/7–8/9 constricted by septum. Length ratio anteseptale: postseptale 1: 2.5 preclitellarly, postseptale tapers gradually into efferent duct, with small terminal vesicle ( Figs. 2D–E View FIGURE 2 ). Dorsal blood vessel from VI ( Figs. 2A View FIGURE 2 , 5A View FIGURE 5 ). Coelomocytes about 40–53 µm long in vivo, flat, cytoplasm pale, margin with indefinite contour in top view, but spindle-shaped in side view ( Fig. 2C View FIGURE 2 ), the length measurable only in side view. Oesophageal appendages in V, well developed without canal in IV ( Figs. 2A View FIGURE 2 , 5A View FIGURE 5 ). Chloragocytes yellowish-brown, about 15 µm long in vivo. Midgut pars tumida XIII–XIV (occupying 2 segments) ( Fig. 2B View FIGURE 2 ). Sperm funnel small, barrel-shaped, 65–90 µm long in vivo (53–62 µm fixed), about 1.2–2 times longer than wide, collar narrower than funnel body ( Figs. 2F–G View FIGURE 2 , 5C View FIGURE 5 ). Sperm duct 6 µm thick in vivo. Spermatozoa 75–90 µm, heads 30–38 µm long in vivo (52–60 µm and 17–23 µm, fixed). Seminal vesicle absent. Male copulatory organs small, widely separate ventro-laterally, no glandular body or bursa, only the pore surrounded by small inconspicuous glands ( Figs. 2H,I View FIGURE 2 ). Spermathecae small, free, confined to V. The short ectal duct (20–30 µm long 20–23 µm wide in vivo) has a small gland ( Figs. 2L,M View FIGURE 2 ). In welldeveloped specimens the duct widens slightly into a narrow dilation of ampulla (25–28 µm wide). After the dilation the connecting tube (about 60 µm long and 20 µm wide) ends in an elongated globular ental reservoir (35–50 µm long, 25–35 µm wide in vivo) in V. The surface of the reservoir is granulated, sperm is visible only in the ampullar dilation and the connecting tube ( Figs. 2J–L View FIGURE 2 , 5B View FIGURE 5 ).
Distribution and habitat. In Korea, Mt. Hallasan, above 1306 m elevation, at site 1–7.
Differential diagnosis. Among the previously described 12 small Achaeta species without pyriform glands and the spermathecae confined to segments V–VI, none has the origin of the dorsal vessel from VI; moreover, secondary pharyngeal glands are, with one exception ( A. hanagarthi Schmelz, 2008 ), present, and the spermathecal opening is without a small gland. In A. camerani (Cognetti, 1899) the dorsal blood vessel originates in VIII and in the all other species in VII. A. brevivasa Graefe, 1980 , A. diddeni Graefe, 2007 , A. hallensis Möller,1976 , A. antefolliculata Dózsa-Farkas & Boros, 2005 , A. afolliculata S esma & Dózsa-Farkas, 1993 and A. singularis Schmelz, 2008 have two pairs of preclitellar nephridia, further A. hanagarthi and A. paranensis Schmelz, 2008 have only one pair. Only A. pannonica Graefe, 1989 , A. iberica Graefe, 1989 and A. etrusca Rota, 1995 similarly to A. koreana sp. n. have three pairs of preclitellar nephridia, as in the new species. But in A. pannonica all reproductive organs except the spermathecae are shifted one segment forward, so the male pores are in XI, while in the new species, in A. iberica and in A. etrusca they are in the usual position in XII. A. etrusca differs from A. koreana sp. n. by his paired knob-like cutaneous gland structures dorsolateral in II–VI and by the shorter spermatozoa (40 µm long, sperm heads 15 µm ( Rota 2015) vs. 75–90 µm and 30–38 µm, respectively), moreover the penial bulbs are compact. Finally, in A. iberica , hyalocytes of the clitellar glands are lined in 4 distinct longitudinal rows.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |