
Vampyrodes major Allen, 1908
|
publication ID |
https://doi.org/10.1206/3710.2 |
|
persistent identifier |
https://treatment.plazi.org/id/03D18783-E039-0862-50B3-FD3C40A0E52C |
|
treatment provided by |
Carolina |
|
scientific name |
Vampyrodes major Allen, 1908 |
| status |
|
Vampyrodes major Allen, 1908 View in CoL
Great Stripe-faced Bat
Figures 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10
Vampyrodes major Allen, 1908: 38 View in CoL ; type locality “San Pablo, Isthmus of Panama,” Canal Zone , Panama .
TYPE MATERIAL: The holotype MCZ 6756, an adult female, preserved in alcohol with the skull removed and cleaned, was collected by Allen Lesley at San Pablo, Isthmus of Panama, Canal Zone, Panama. The body and skull are in good condition. The type locality is now covered by the waters of Gatún Lake ( Goldman, 1920) .
DISTRIBUTION: Vampyrodes major is known from southern Mexico ( Chiapas and Oaxaca), Belize, Guatemala, Honduras, Nicaragua, Honduras, Costa Rica, Panama, western Colombia and Ecuador (fig. 6). V. major is expected to occur in El Salvador, but has not been reported yet ( Burt and Stirton, 1961; Owen et al., 1991).
EMENDED DIAGNOSIS: Dorsal fur is dark brown, 9–10 mm long; three genal vibrissae present; inverted V-shaped posterior margin of the uropatagium; uropatagium fringed with long (> 2 mm), dense hair along trailing edge; metacarpal III shorter than metacarpal V; rostrum broad and robust; parietal foramina close to nuchal crest; absent or weakly developed anterior notch in the nasals; well-developed groove present between the occipital condyle and paracondylar process; paraoccipital processes moderately developed; perikymata present on all upper and lower teeth; I1 slender and bilobed but appears single lobed in older individuals with worn teeth; M1 postentoconule absent or poorly developed; M2 parastyle well developed; M2 postentoconule well developed; lower incisors small and bilobed; lingual accessory cuspule present on p4; cuspule on m1 paracristid present; cuspule on m2 paracristid sometimes present and sometime absent.
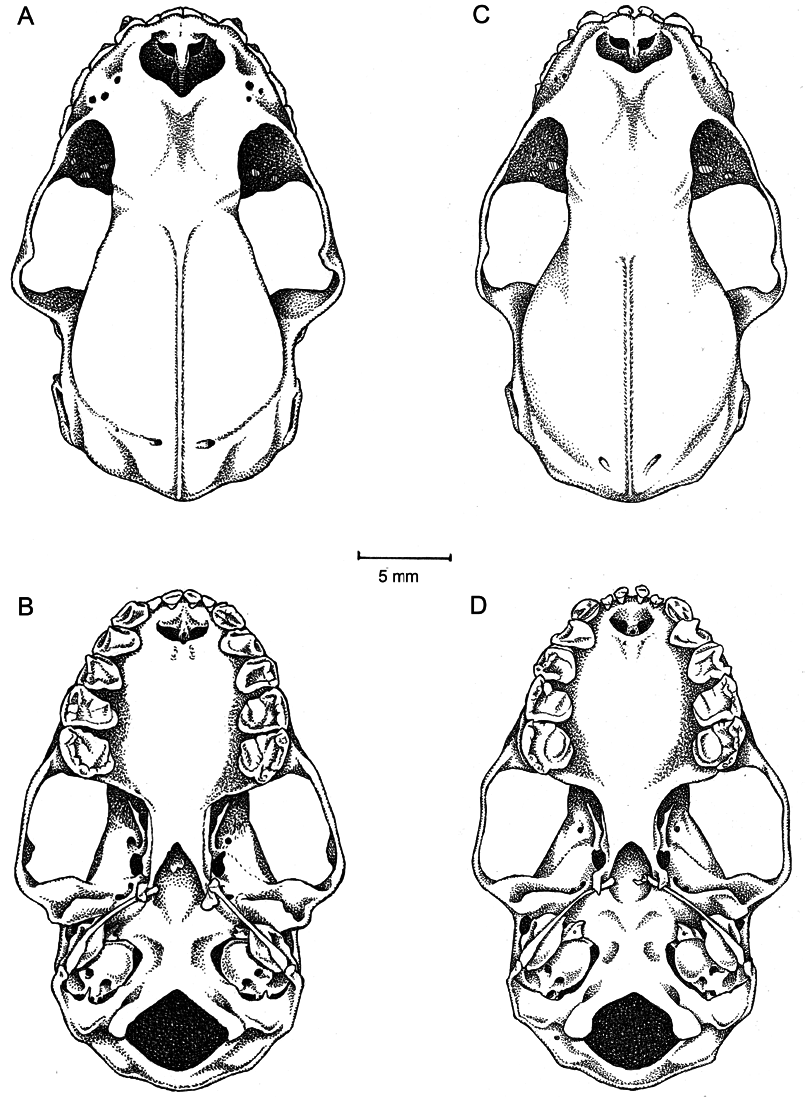
DESCRIPTION AND COMPARISONS: A medium-sized Vampyrodes (FA 51.36–58.64 mm; GLS 26.26–29.06 mm; CCL 23.09–25.71 mm; table 5). All linear measurements of V. major show a small overlap with those of V. caraccioli with V. major being the larger of the two species (tables cioli (USNM 405129; male); the stylohyal was reconstructed from USNM
582872. Lateral views of the skulls and lower jaw of ( B) V. major (FMNH
127114; male).
4–5). The dorsal pelage in V. major is long ( 9–10 mm) and brown, with individual hairs bicolored with darker tips. Compared with V. caraccioli , the pelage tends to slightly darker and the hairs longer ( 7–9 mm in V. caraccioli ). The ventral pelage is similar but slighty darker than in the latter species, with individuals hairs tricolored, with a basal pale brownish band that makes up some 70% to 80% of the total length of each hair, a short dark brown (~ 10% of the total length of each hair) subterminal band, and a tiny pale brownish terminal band. The uropatagium in V. major has an inverted V-shaped posterior margin with dense and long hair (> 2 mm) along the trailing edge (U-shaped posterior margin, with dense and short hair (< 2 mm) along the trailing edge in V. caraccioli ). The width of uropatagium in V. major is 6–10 mm long at midline (in V. caraccioli the uropatagium tends to be slightly shorter, 5–9 mm). The proximal half of forearm in V. major is covered with dense, short hair. Metacarpal III is shorter than metacarpal V in V. major (metacarpal III is longer than metacarpal V in V. caraccioli ). The plagiopatagium inserts onto the tarsal bones in both species. Three genal vibrissae are present in V. major (two genal vibrissae are present in V. caraccioli ). V. major has six vibrissae surrounding the margin of the noseleaf in a single array; two vibrissae on each side of upper lip below the vibrissae surrounding the noseleaf; four submental vibrissae on each side of chin; two interramal vibrissae. The noseleaf is longer than wide and the inferior border of nasal horseshoe is completely free of upper lip.
The skull of V. major has a broad and robust rostrum (slender in V. caraccioli ) and the anterior notch in the nasals is absent or weakly developed (well developed in V. caraccioli ). Two infraorbital foramina are usually present (multiple infraorbital foramina [> 4] are present on one specimen examined: USNM 309833). The parietal foramina are close to the nuchal crest (FMNH 127114; fig. 10B), whereas they are located much further from the nuchal crest in V. caraccioli (USNM 405129; fig. 10A). The groove between the occipital condyle and paracondylar process is well developed (FMNH 58263; fig. 10B); in comparison, in V. caraccioli the groove between the occipital condyle and paracondylar process is weakly developed (USNM 405129; FMNH 139776; fig. 10A). The paraoccipital processes are moderately developed in V. major (well developed in V. caraccioli ).
Perikymata are present in all upper and lower teeth in V. major . The upper inner incisors are slender; both outer and inner incisors are bilobate (FMNH 127114; USNM 539812), but appear single lobed in older individuals with worn teeth (AMNH 186381). Within comparison, in V. caraccioli , the upper inner incisors are mesiodistally broad. P3 is more than half the size of P4, and there are two stylar cuspules present on posterior cristid of P 4 in V. major . The M1 parastyle is absent, but both a mesostyle and metastyle are present, and a stylar cuspule is present on the labial cingulum of the M1 metacone. A sulcus is present on the posterior cristid of the M1 paracone, the protocone is well developed, and the postentoconule is absent or poorly developed on M1. On M2 the parastyle is well developed (FMNH 127114) (absent or poorly developed in V. caraccioli ), the labial cingulum on the paracone is absent or poorly developed, and the postentoconule is well developed (absent or poorly developed in V. caraccioli ). The lower incisors are small and bilobed (robust and bilobed in V. caraccioli ). A p4 lingual accessory cuspule is present. A cuspule on the m1 paracristid is present in V. major (absent in V. caraccioli ). A cuspule on the m2 paracristid is sometimes present (AMNH 186381) and sometimes absent (USNM 314717) in V. major . In V. caraccioli , this cuspule is always absent.
NATURAL HISTORY: The natural history of Vampyrodes major has been extensively studied (under the name V. caraccioli ) Barro Colorado Island, Panama ( Bonaccorso, 1979; Giannini and Kalko, 2004). V. major is a frugivorous bat that has been reported to take fruits/infrutescences of 13 species representing five genera in four families: Spondias mombin , Spondias radlkoferi (Anacardiaceae) ; Calophyllum longifolium (Clusiaceae) ; Ficus dugandii , F. insipida , F. maxima , F. obtusifolia , F. pertusa , F. trigonata , F. yoponensis , F. sp. and Poulsenia armata (Moraceae) , and Piper sp. (Piperaceae) ( Bonaccorso, 1979; Morrison, 1980; Handley et al., 1991; Kalko et al., 1996; Medellín and Gaona, 1999; Wendeln et al., 2000; Giannini and Kalko, 2004).
Four species of ectoparasites have been obtained from Panamanian specimens of Vampyrodes major: Chirnyssoides caparti ( Sarcoptidae ), Parichoronyssus sp. (Macronyssidae) , Strebla vespertilionis (Streblidae) , and Periglischrus iheringi (Spinturnicidae) ( Furman, 1966; Wenzel et al., 1966).
Like Vampyrodes caraccioli View in CoL , V. major View in CoL has a litter size of one ( Davis et al., 1964; Jones, 1964; Jones et al., 1971). Reproductive data suggested seasonal polyestry; pregnant females have been captured in all months but in December ( Davis et al., 1964; Jones, 1964; Jones et al., 1971; Valdez and LaVal, 1971; Fleming et al., 1972; Thomas, 1972; Bonaccorso, 1979; Estrada and Coates-Estrada, 2001). Lactating females have been captured from January to September ( Davis et al., 1964; Thomas, 1972; Bonaccorso, 1979).
KARYOLOGY: Vampyrodes major View in CoL has a similar karyotype of V. caraccioli View in CoL (2 n = 30, FN = 56).
The X chromosome is subtelocentric and the Y chromosome is submetacentric ( Baker, 1973).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Vampyrodes major Allen, 1908
| Velazco, PaúL M. & Simmons, Nancy B. 2011 |
Vampyrodes major
| Allen, G. M. 1908: 38 |