
Dracoderes snufkini, Yamasaki, Hiroshi, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3980.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:1E024E35-8591-45AD-B23A-7A21F615A0EF |
|
DOI |
https://doi.org/10.5281/zenodo.6095454 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF87B4-FFF6-FFC2-FF1B-FB52305FF85A |
|
treatment provided by |
Plazi |
|
scientific name |
Dracoderes snufkini |
| status |
sp. nov. |
Dracoderes snufkini sp. nov.
[New Japanese name: Ryukyu tatsutogekawa] ( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ; Tables 3 View TABLE 3 , 4 View TABLE 4 )
Dracoderes sp. 1. Sørensen et al. 2012: 228 –229, fig. 11.
Material examined. Holotype: Adult female (ZIHU-04984), collected 25 January 2014 at station 1 ( Fig. 1 View FIGURE 1 ), mounted in Fluoromount G®. Allotype: Adult male (ZIHU-04985), station 1, mounted in Fluoromount G®. Paratypes: One adult female and one adult male (ZIHU-04986–04987), two adult females and one adult male (RUMF-ZK-00019–00021), one adult female and two adult males ( ZMUC KIN- 000834–000836), all from station 1, all mounted in Fluoromount G®. Additional material: Two male exoskeletons (ZIHU-04988–04989) from specimens for DNA extraction, collected 17 July 2013, station 2 ( Fig. 1 View FIGURE 1 ), mounted in Fluoromount G®. Three males, three females, and two sex-undetermined specimens for SEM, station 1, 25 January 2014, mounted on aluminum stubs stored in the author’s laboratory (University of the Ryukyus, Okinawa, Japan). Sequences: 18S (1594 bp) from ZIHU-04989, GenBank accession number LC 032113 View Materials ; 28S (3249 bp) from ZIHU-04989, LC 032114 View Materials ; COI (629–658 bp) from ZIHU-04988–04989 and one specimen for which exoskeleton was lost, LC 032115 View Materials – LC 032117 View Materials .
Type locality. Oura Bay, Okinawa, Japan (26°31.863' N, 128°3.179' E).
Diagnosis. Dracoderes with lateroventral tubules on segments 2 and 5; thick and plump middorsal spines on segments 2 and 9; thick and plump paradorsal spines on segments 3–8, alternately laterally displaced; ventral primary pectinate fringe on segment 1 with long, wide, and conspicuous tips.
Etymology. The specific epithet snufkini is after the Diving Team Snack Snufkin, which advertises the beauty, greatness, and importance of nature in Oura Bay, Okinawa, the type locality of the new species. ‘Snufkin’ is also the nickname for Mr Nishihira, a delegate on the team. The team also cooperated with the 1st Umisawa Kai (Field workshop for young marine biologists) held in Oura Bay from 24–28 January 2014, and helped collecting the new species.
Of the Japanese name, ‘Ryukyu’ is from the Ryukyu Islands where the species was found, and ‘tatsutogekawa’ is from the Japanese name for the genus Dracoderes proposed in Murakami (2005).
Description. Adult with head, neck, and eleven trunk segments ( Figs 2 View FIGURE 2 A, B, 3A, 4A). See Table 3 View TABLE 3 for measurements. Table 4 View TABLE 4 indicates the positions of cuticular structures.
Head consists of retractable mouth cone and introvert. Mouth cone with inner oral styles and outer oral styles. Introvert with spinoscalids and at least nine trichoscalids. Exact number and arrangement of inner oral styles, outer oral styles, and spinoscalids not determined.
Neck with four dorsal and five ventral placids ( Figs 2 View FIGURE 2 A, B, 3B, E, 4B, C, E). Midventral and midlateral placids slightly wider than others. Ventral placids close together, dorsal placids at intervals occupied by cuticular folding ( Figs 2 View FIGURE 2 A, B, 3B, E, 4B). Surface of placids anteriorly covered with fine hairs, surface of posterior part smooth, lacking any cuticular structures ( Fig. 4 View FIGURE 4 B).
Trunk with 11 segments; segment 1 consists of complete cuticular ring; segments 2–11 consist of one tergal and two sternal plates ( Figs 2 View FIGURE 2 A, B, 3A). Thickened cuticle forms pachycyclus at anterior margin of all segments ( Figs 2 View FIGURE 2 A, B, 3A–E). Middorsal acicular spines on segments 2 and 9 ( Figs 2 View FIGURE 2 A, C, 3D, E, 4F, 5B). Paradorsal acicular spines on segments 3–8, laterally displaced alternately right and left ( Figs 2 View FIGURE 2 A, C, 3D, E, 4F, 5A, B). All dorsal spines short, plump and pointed. Funnel-shaped subcuticular structures visible at base of each dorsal acicular spine under LM observation ( Figs 2 View FIGURE 2 A, C, 3D, E). Pair of lateral accessory tubules on segment 8, difficult to find under LM but visible with SEM ( Fig. 5 View FIGURE 5 B–D). Pair of lateroventral tubules on segments 2 and 5 ( Figs 2 View FIGURE 2 B, 3B, C, 4C, 5A). Pair of lateroventral acicular spines on segments 6–9 ( Figs 2 View FIGURE 2 B, D, 3C, 5A–E). All sensory spots rounded, with collar of papillae around single central pore ( Fig. 4 View FIGURE 4 D). Cuticular hairs on posterior part of segment 1, tergal plates of segments 2–9, and posterior part of sternal plate of segments 2–9 ( Figs 2 View FIGURE 2 A, B, 4C, E–G, 5A–E). No secondary pectinate fringe or glandular cells were observed.
Segment 1 shows conspicuous primary pectinate fringe with long, wide spinous processes on ventral side ( Figs 2 View FIGURE 2 B, 3B, 4C, G), each spinous process observable by both LM and SEM. Dorsal to lateral primary pectinate fringe with short, densely arranged tips, difficult to identify individually with LM ( Figs 2 View FIGURE 2 A, B, 3E, 4E, G). Pair of sensory spots subdorsally, laterodorsally, and midlaterally; two pairs of sensory spots ventromedially ( Figs 2 View FIGURE 2 A, B, 3B, E, 4C, E–G).
Segment 2 with middorsal acicular spine and lateroventral tubules ( Figs 2 View FIGURE 2 A, B, 3B, E, 4C, F). Pairs of sensory spots in ventrolateral and ventromedial positions ( Figs 2 View FIGURE 2 B, 3B). Two pairs of sensory spots in laterodorsal position ( Figs 2 View FIGURE 2 A, 3E, 4F, G). Primary pectinate fringe with short, densely arranged tips ventromedially to midventrally. Middorsal and lateroventral parts of fringe show long, thin tips and area of short, densely arranged tips, respectively ( Fig. 4 View FIGURE 4 G).
Segment 3 with paradorsal acicular spine located to the left or to the right ( Figs 2 View FIGURE 2 A, 3D, E, 4F). Perispinal paradorsal sensory spots present ( Figs 2 View FIGURE 2 A, 3D, E). Additional sensory spots present in laterodorsal, midlateral and ventrolateral positions ( Figs 2 View FIGURE 2 A, B, 3B, E, 4F, G). Primary pectinate fringe of sternal plates with short, densely arranged tips. Fringe of tergal plate similar to that of preceding segment ( Fig. 4 View FIGURE 4 G).
Segment 4 with paradorsal acicular spine, on the opposite side of that on the preceding segment ( Figs 2 View FIGURE 2 A, 3D, E, 4F). Perispinal paradorsal sensory spots present ( Figs 2 View FIGURE 2 A, 3D, E). Additional sensory spots in laterodorsal, ventrolateral, and ventromedial positions ( Figs 2 View FIGURE 2 A, B, 3B, E, 4F). Primary pectinate fringe similar to than the fringe on the preceding segment.
Segment 5 similar to segment 3, except for presence of lateroventral tubules ( Figs 2 View FIGURE 2 B, 3C, 4F, 5A).
Segment 6 with paradorsal acicular spine and lateroventral acicular spines ( Figs 2 View FIGURE 2 A, B, 3C, D, 5A). Perispinal subdorsal sensory spots present ( Figs 2 View FIGURE 2 A, 3D). Additional sensory spots in laterodorsal, midlateral, ventrolateral, and ventromedial positions ( Figs 2 View FIGURE 2 A, B, 5A). Primary pectinate fringe on this and following four segments with short, densely arranged tips on tergal and sternal plates.
Segment 7 similar to segment 6 except for absence of sensory spots in ventromedial position ( Fig. 5 View FIGURE 5 A–C).
Segment 8 similar to segment 6 except for the presence of lateral accessary tubules, difficult to identify with LM, but visible with SEM ( Fig. 5 View FIGURE 5 B–D).
Segment 9 with middorsal acicular spine and lateroventral acicular spines ( Figs 2 View FIGURE 2 A–D, 3C, D, G, 5B–E). Sensory spots in subdorsal, laterodorsal, midlateral, ventrolateral, and ventromedial positions ( Figs 2 View FIGURE 2 A–D, 3C, D, 5B–D). Nephridial pores small, posteriorly directed opening with a few papillae in lateral accessory position ( Fig. 5 View FIGURE 5 D).
Segment 10 lacking acicular spine and tubule. Sensory spots in subdorsal, laterodorsal, and ventrolateral positions ( Figs 2 View FIGURE 2 A–D, 3C, G, 5E).
Segment 11 with lateral terminal spines ( Figs 2 View FIGURE 2 A–D, 3A, C, F, G, 4A, 5E). Lateral terminal accessory spines lacking in both sexes. Females with slit-like gonopore openings in lateroventral position ( Fig. 3 View FIGURE 3 G). Males with three pairs of penile spines ( Fig. 2 View FIGURE 2 D, 3F, 5E). Two pairs of sensory spots subdorsally; one in central part of segment, another at tip of tergal extension ( Fig. 2 View FIGURE 2 A, C). Additional sensory spots in ventrolateral position ( Figs 2 View FIGURE 2 B, D, 3G, 5E). Primary pectinate fringe with short, densely arranged tips laterally and ventrally ( Fig. 5 View FIGURE 5 E). Posterior end with triangular tergal extension dorsally. Ventral side of tergal extension covered densely with minute hairs ( Fig. 5 View FIGURE 5 E).
Remarks. Dracoderes snufkini sp. nov. differs markedly from congeners in the shape of the primary pectinate fringe on the ventral side of segment 1; it has a primary pectinate fringe strongly developed, i.e., the tips of the pectination are conspicuously long and wide on the ventral side on segment 1, whereas in congeners the primary pectinate fringe on segment 1 consists of relatively narrow, short inconspicuous tips ( Higgins & Shirayama 1990; Adrianov & Malakhov 1999; Sørensen et al. 2012; Thomsen et al. 2013). Dracoderes snufkini also differs from congeners (except D. orientalis ) in lacking lateroventral tubules on segment 10, and from D. orientalis in having lateroventral tubules on segments 2 and 5 ( Adrianov & Malakhov 1999).
Dracoderes snufkini is very similar to Dracoderes sp. 1 in Sørensen et al. (2012). Similarities include 1) the same numbers and positions of spines and tubules, 2) short, plump dorsal spines, 3) similar width/length of trunk, trunk segments and spines ( Table 3 View TABLE 3 ), and 4) a strongly developed primary pectinate fringe on the ventral side on segment 1. Sørensen et al. (2012) described Dracoderes sp. 1 as having a strongly developed primary pectinate fringe on segment 2, but this character was not found by me in his specimens. I assume the posterior margin of segment 1 was mistaken as the posterior margin of segment 2 ( Sørensen et al. 2012: Fig. 11C). In addition, Dracoderes sp. 1 was found at Kabira Bay, Ishigaki Island, Okinawa, very close to one of the localities where D. snufkini was collected ( Fig. 1 View FIGURE 1 ). These morphological and geographic similarities indicate that specimens referred to as Dracoderes sp. 1 in Sørensen et al. (2012) are conspecific with D. snufkini .
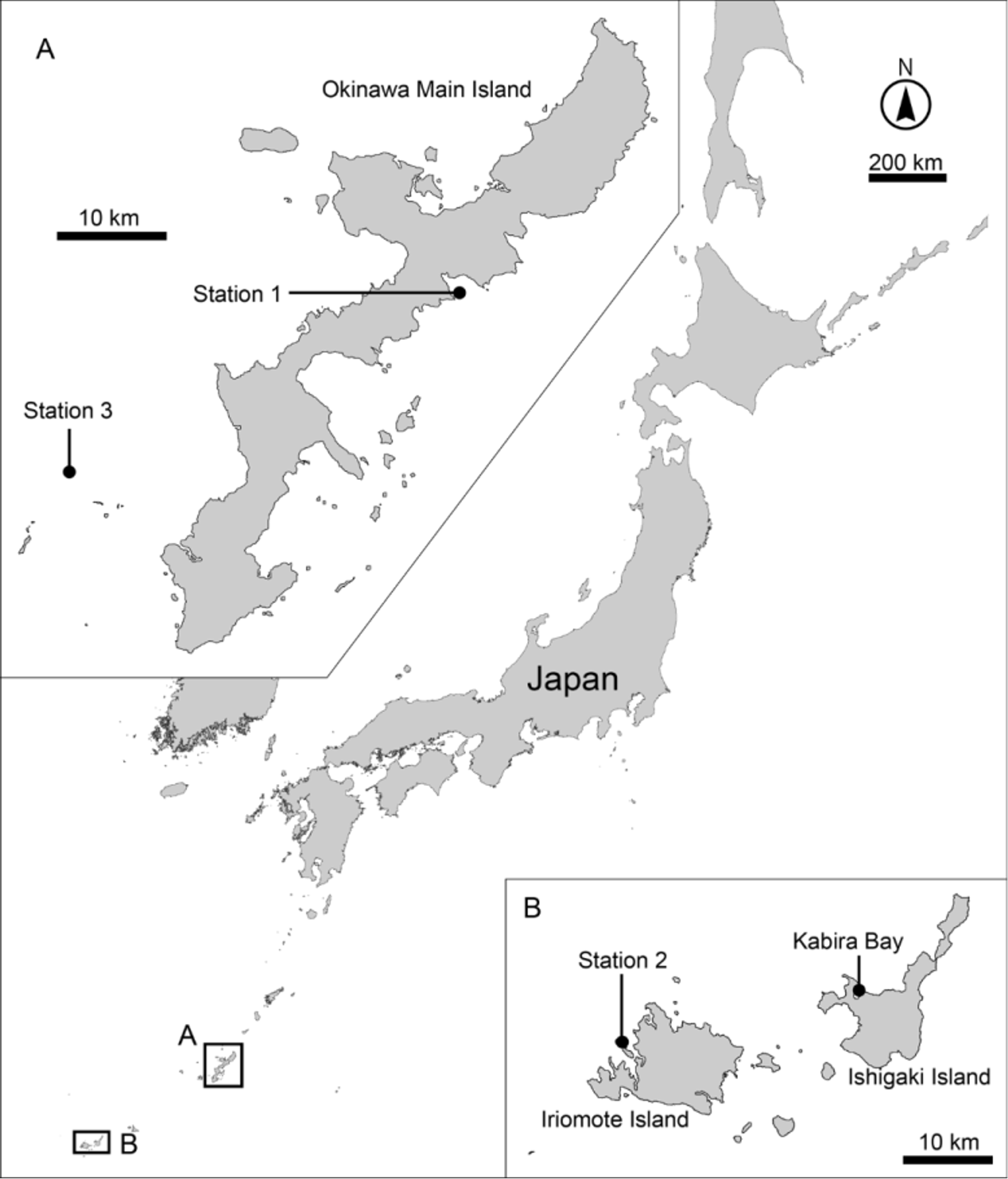




TABLE 3. Measurements for adult Dracoderes snufkini sp. nov. (in micrometers). Measurements are respectively given for the type specimens from Okinawa Main Island, specimens from Ishigaki Island deposited as Dracoderes sp. in the Zoological Museum, Natural History Museum of Denmark, and specimens in total. Columns N and SD indicate sample size and standard deviation, respectively. Abbreviations: (ac), acicular spine; LA, length of lateral accessory tubule; LTS, length of lateral terminal spine; LV, length of lateroventral spine / tubule; MD, length of middorsal spine; MSW, maximum sternal width; PD, length of paradorsal spine; S, segment length; SW, standard width; TL, trunk length.
| Character | Okinawa Main Island N Range Mean | SD | Ishigaki Island N Range | Mean | SD | total N Range | Mean | SD |
|---|---|---|---|---|---|---|---|---|
| TL MSW-6 MSW-6/TL | 10 249–283 267 10 67–72 69 10 24.0–28.5% 26.0% | 10.70 1.70 1.47% | 13 206–280 11 62–75 11 26.2–28.4% | 252 70 27.1% | 23.29 3.61 0.8% | 23 206–283 21 62–75 21 24.0–28.5% | 259 70 26.6% | 20.09 2.82 1.27% |
| SW-10 SW-10/TL | 10 53–60 56 10 19.5–23.4% 21.1% | 2.26 1.33% | 11 54–63 11 19.9–24.8% | 57 22.0% | 2.57 1.36% | 21 53–63 21 19.5–24.8% | 57 21.6% | 2.38 1.39% |
| S1 S2 | 10 28–39 36 10 19–23 21 | 2.81 1.28 | 11 32–37 11 19–23 | 35 21 | 1.99 1.33 | 21 28–39 21 19–23 | 35 21 | 2.36 1.28 |
| S3 S4 | 10 21–25 23 10 24–27 25 | 1.49 1.12 | 11 21–28 11 22–29 | 23 25 | 1.96 2.24 | 21 21–28 21 22–29 | 23 25 | 1.74 1.76 |
| S5 S6 | 10 26–30 28 10 27–30 29 | 1.33 1.23 | 11 23–33 11 23–33 | 28 29 | 2.36 2.65 | 21 23–33 21 23–33 | 28 29 | 1.9 2.05 |
| S7 S8 S9 | 10 27–34 29 10 24–31 28 10 23–30 27 | 1.85 1.90 2.96 | 11 26–32 11 28–35 11 28–34 | 30 31 31 | 1.84 1.96 1.57 | 21 26–34 21 24–35 21 23–34 | 30 30 29 | 1.83 2.44 3.27 |
| S10 S11 | 10 20–26 24 10 26–30 27 | 2.07 1.30 | 11 24–31 11 26–30 | 27 28 | 2.00 1.13 | 21 20–31 21 26–30 | 25 27 | 2.58 1.19 |
| MD2 (ac) PD3 (ac) | 10 22–25 23 10 24–30 27 | 1.22 1.42 | 13 23–28 13 25–33 | 26 29 | 1.96 2.57 | 23 22–28 23 24–33 | 24 28 | 1.99 2.28 |
| PD4 (ac) PD5 (ac) PD6 (ac) | 10 27–31 29 10 29–31 30 10 27–31 29 | 1.18 0.77 1.38 | 13 27–34 13 28–37 13 26–35 | 31 32 31 | 2.18 2.48 2.40 | 23 27–34 23 28–37 23 26–35 | 30 31 30 | 2.07 2.26 2.15 |
| PD7 (ac) PD8 (ac) | 10 26–33 28 10 22–27 25 | 2.10 1.52 | 13 28–35 11 27–31 | 31 29 | 2.04 1.22 | 23 26–35 21 22–31 | 30 27 | 2.6 2.57 |
| MD9 (ac) LV2 (tu) | 10 22–30 27 10 10–15 12 | 2.74 1.44 | 13 25–33 2 9–10 | 30 10 | 2.8 0.50 | 23 22–33 12 9–15 | 28 12 | 3.09 1.71 |
| LV5 (tu) LV6 (ac) | 10 14–17 16 10 22–27 24 | 1.13 1.31 | 3 15–19 11 22–28 | 17 26 | 2.29 1.86 | 13 14–19 21 22–28 | 16 25 | 1.45 1.81 |
| LV7 (ac) LV8 (ac) | 10 25–28 26 10 23–27 25 | 0.83 1.38 | 11 27–31 11 24–30 | 29 27 | 1.17 1.90 | 21 25–31 21 23–30 | 27 26 | 1.68 1.98 |
| LA8 (tu) LV9 (ac) LTS | 10 10–14 15 10 23–26 25 9 187–206 195 | 1.24 0.85 6.57 | 0 - 11 26–30 13 176–214 | - 27 195 | - 1.55 11.1 | 10 10–14 21 23–30 22 176–214 | 12 26 195 | 1.24 1.66 9.32 |
| LTS/TL | 9 67.8–79.2% 73.3% | 3.52% | 13 68.8–90.6% | 78.0% | 7.51% | 22 67.8–90.6% | 76.1% | 6.53% |
TABLE 4. Summary of locations of cuticular structures, tubules, and spines in Dracoderes snufkini sp. nov. Abbreviations: ac, acicular spine; LA, lateral accessory; LD, laterodorsal; lts, lateral terminal spine; LV, lateroventral; (m), male condition of sexually dimorphic character; MD, middorsal; ML, midlateral; nep, nephridial opening; PD, paradorsal; pe, penile spine; per, located perispinally; SD, subdorsal; ss, sensory spot; tu, tubule; VL, ventrolateral; VM, ventromedial.
| Position | MD | PD | SD | LD | ML | LA | LV | VL | VM |
|---|---|---|---|---|---|---|---|---|---|
| segment | |||||||||
| 1 | ss | ss | ss | ss, ss | |||||
| 2 | ac | ss, ss | tu | ss | ss | ||||
| 3 | ac, ss (per) | ss | ss | ss | |||||
| 4 | ac, ss (per) | ss | ss | ss | |||||
| 5 | ac, ss (per) | ss | ss | tu | ss | ||||
| 6 | ac | ss (per) | ss | ss | ac | ss | ss | ||
| 7 | ac | ss (per) | ss | ss | ac | ss | |||
| 8 | ac | ss (per) | ss | tu | ac | ss | ss | ||
| 9 | ac | ss | ss | ss | nep | ac | ss | ss | |
| 10 | ss | ss | ss | ||||||
| 11 | ss, ss | pe (m) | lts | ss |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dracoderes snufkini
| Yamasaki, Hiroshi 2015 |
Dracoderes
| Sorensen 2012: 228 |