
Calotriton, GRAY, 1858
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00197.x |
|
persistent identifier |
https://treatment.plazi.org/id/03CE8B72-BB04-FA22-FC19-85DE7190FE84 |
|
treatment provided by |
Diego |
|
scientific name |
Calotriton |
| status |
|
GENUS CALOTRITON GRAY, 1858 View in CoL
Calotriton Gray, 1858: 139 View in CoL
Type species: Hemitriton punctulatus Dugès, 1852 by monotypy (= Calotriton asper ( Dugès, 1852)) Hemitriton Dugès, 1852: 266 . Type species not designated. Name already occupied by Hemitriton Hoeven, 1833: 305 .
Remarks
The use of Hemitriton Dugès, 1852 for the Western brook newts pre-dates Calotriton Gray, 1858 , but it is already occupied by Hemitriton Hoeven, 1833 , so Calotriton is the first available name for the group.
Diagnosis
Small- to medium-sized newts (70–167 mm including tail). Fronto-squamosal arch bony. Palatine teeth in two series in contact anteriorly, diverging posteriorly, therefore forming an inverted Y-shaped figure. The series starts on a line with the choanae. Tongue small, elliptical, the sides slightly free. Head depressed, longer than wider, fairly robust (especially males), with the intranasal cavity elongated and well developed. The greatest head width is at the posterior corners of eyes. Prominent swellings on the posterior sides of the head (see detail in Fig. 10A, C View Figure 10 ). Upper jaw overhanging with well-developed lateral upper labial folds extending over the edge of the lower jaw. Paratoid glands absent. Gular fold present. Skin covered with tubercles bearing horny tips, more or less strongly tuberculate above, less tuberculate or completely smooth beneath. Limbs moderate, four fingers and five toes, all depressed and free, with the tips covered by a black corneous nail-like sheath. No distinct spurs on male or female hind legs. Body rounded or slightly depressed, absence of cutaneous dorsal and caudal crest even during the breeding season. Tail ending in an obtuse point and about as long as head and body (longer in females) and compressed from side, deeper in males. Lungs very reduced. Cloacal swelling rounded and with a vertical slit in males, and conical or cylindrical, directed backwards, in females. Caudal capture of female to facilitate direct transfer of spermatophores.
Species
Calotriton asper ( Dugès, 1852) View in CoL and C. arnoldi View in CoL sp. nov.
Distribution
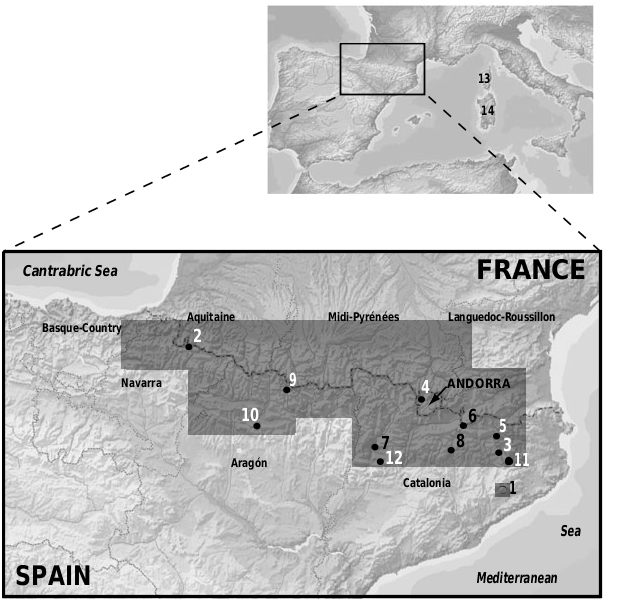
The distribution range includes most of the Pyrenees, the Massif of El Montseny and some intermediate areas (see shadowed area in Fig. 1 View Figure 1 ).
CALOTRITON ASPER ( DUGÈS, 1852) View in CoL
( FIGS 4F–I, P–R View Figure 4 , 10K–L View Figure 10 )
Triton glacialis Philippe 1847: 20 View in CoL (nomen nudum) Hemitriton repandus Duméril & Valenciennes, 1852 . In Dugès, 1852: 256 (nomen nudum)
Hemitriton cinereus Dugès, 1852: 263 View in CoL , pl. 1, figs 14, 15.
Hemitriton rugosus Dugès, 1852: 264 View in CoL , pl. 1, figs 16, 17.
Hemitriton puncticulatus Duméril 1852 . In Dugès, 1852: 265 (nomen nudum)
Hemitriton punctulatus Dugès, 1852: 265 View in CoL , pl. 1, fig. 18.
Hemitriton asper Dugès, 1852: 266 View in CoL (nomen conservandum)
Hemitriton bibronii Dugès, 1852: 266 View in CoL , pl. 1, figs 19, 20.
Triton Pyrenaeus Duméril, Bibron & Duméril, 1854: 139 View in CoL .
Triton rugosus Duméril, Bibron & Duméril, 1854: 150 View in CoL . Triton cinereus Duméril, Bibron & Duméril, 1854: 151 View in CoL . Triton repandus Duméril, Bibron & Duméril, 1854: 151 View in CoL , ATLAS, pl. 106, fig. 2.
Triton puncticulatus Duméril, Bibron & Duméril, 1854: 152 View in CoL . Incorrect spelling of punctulatus .
Triton Bibroni View in CoL (incorrect spelling of bibronii Dugès ). Duméril, Bibron & Duméril, 1854: 153 (nomen nudum).
Calotriton punctulatus Gray, 1858: 139 View in CoL .
Euproctus Pyrenaeus Lataste, 1878: 495 View in CoL .
Molge aspera Boulenger, 1882: 24 View in CoL ; Maluquer, 1916: 58. Euproctus aspera Camerano, 1885: 419 View in CoL (lapsus).
Euproctus asper var. rugosa Bedriaga, 1895: 150 View in CoL .
Triton asper Wolterstoff, 1900: 31 View in CoL ; Wolterstoff, 1902: 12.
Molge bolivari Bosca, 1918: 58 View in CoL .
Euproctus asper Wolterstoff, 1925: 61 View in CoL ; Wolterstoff & Herre, 1935: 224; Thorn, 1968: 253.
Euproctus asper var. peyreladensis Wolterstoff, 1925: 71 .
Euproctus asper View in CoL var. d’Onceti Wolterstoff, 1925: 66.
Euproctus asper var. castelmouliensis Wolterstoff, 1925: 66 View in CoL ; Mertens & Wermuth, 1960: 17.
Euproctus (Hemitriton) asper Wolterstoff, 1925: 296 View in CoL .
Euproctus asper asper Mertens & Muller, 1928: 9 View in CoL ; Mertens & Wermuth, 1960: 17.
Euproctus asper castelmouliensis Mertens & Muller, 1940: 8 View in CoL ; Mertens & Wermuth, 1960: 17.
Diagnosis
Maximum total length of preserved males included in our study 127.5 mm, females 143 mm. Measurements for 14 other morphometric characters are presented in Tables 6, 8 for males and females, respectively. First 3–4 caudosacral vertebrae with prominent transverse processes at approximately 90° to the vertebral axis ( Fig. 4F–I, P–R View Figure 4 ). This character is very clear in adult male specimens ( Fig. 4F–H, P–R View Figure 4 ) and less evident in females and juvenile specimens ( Fig. 4I View Figure 4 , our pers. observ.). Skin usually very rough, covered with a high proportion of tubercles bearing horny tips (including the venter and underside of the tail; see Fig. 10K, L View Figure 10 ). Usually greyish, olive, or blackish above, uniform or with light yellowish spots, which often are confluent into a broad vertebral line, especially in subadults and younger adults. Venter opaque with a yellow, orange or red central band usually unspotted with large dark markings at sides or with few spots. Cloacal swelling bulbous-conical in females, usually relatively broad at the base (frequently broader than the tail) ( Fig. 10K, L View Figure 10 ).
Material examined
Museum specimens used in the multivariate morphological analysis are listed in Appendix 1. Specimens included in the X-ray osteological analysis are listed in Appendix 2. Observations on live material include specimens from the whole distribution range of the species (see Fig. 1 View Figure 1 ).
CALOTRITON ARNOLDI SP. NOV.
( FIGS 4A–E, S View Figure 4 , 10A–J View Figure 10 )
Euproctus asper ( Dugès, 1852: 1) View in CoL (part); Llorente et al., 1995: 34; Montori, 1988: 20; Montori & Campeny, 1991: 1; Montori et al., 1997: 106; Montori et al., 2002: 48; Montori & Pascual, 1981: 1; Salvador & García-París, 2001: 35; García-París et al., 2004: 120.
Diagnosis
Similar to Calotriton asper but differs in the following features: mitochondrial DNA sequences different (divergence: cytb, 4.1%; 12S rRNA, 1.2%; 16S rRNA, 0.7%; GenBank accession numbers: DQ092234 View Materials –40, DQ092281 View Materials –82, DQ092299 View Materials -300). First caudosacral vertebrae with short transverse processes that are directed obliquely backwards ( Fig. 4A–E, S View Figure 4 ) (not elongate and at approximately right angles to the vertebral axis as in C. asper ; Fig. 4F–I, P–R View Figure 4 ). In each sex, spiny-tipped tubercles on the dorsum smaller and fewer, so surface generally much smoother than in C. asper ( Fig. 10A–D View Figure 10 ). Tubercles never present in the venter or underside of the tail ( Fig. 10F, I–J View Figure 10 ) (can be present in C. asper Fig. 10K, L View Figure 10 ). Maximum body-size of male and female alcohol-preserved specimens less (maximum females 102.62 mm, maximum males 102.25 mm), compared with 143 mm and 127.5 mm of maximum total length of adult female and male C. asper , respectively. Dorsum dark, chocolatecoloured ( Fig. 10A–D View Figure 10 ), sometimes with light silverygold stippling on the sides. Adult and young C. arnoldi from populations B1 and B2 always uniform dorsally, with a light and very thin brownish-orange stripe from the base to the tip of the tail, sometimes extending further into the body (see Fig. 10D View Figure 10 ). Some adult and young specimens from populations A1–A3, apart from the thin orange stripe mentioned above, also present light greenish or yellowish blotches on the sides of the tail and body ( Fig. 10B View Figure 10 ), these being more obvious when young ( Fig. 10G, H View Figure 10 ). Complete absence in both adults and young of the broad yellow or light-coloured vertebral stripe common in C. asper (especially in young C. asper ). Throat very light, of a pink-ivory colour, largely unspotted or with a lightly dark stippling ( Fig. 10F View Figure 10 ). Venter translucent (follicles can be clearly seen by transparency in adult females), of a light ochre-brown colour, with dark markings of approximately the same colour as the dorsum consisting of dark stippling that are best developed at the sides ( Fig. 10F View Figure 10 ) (belly opaque with large dark spots or blotches usual in C. asper ). Throat posterior to gular region with a relatively large whitish-ivory immaculate blotch (see Fig. 10F View Figure 10 ). Female cloaca cylindrical, relatively narrow and of a contrasted bright reddishorange colour at the tip ( Fig. 10F, I–J View Figure 10 ) (usually bulbous-conical, broader at the base in C. asper ; see Fig. 10K, L View Figure 10 ). When manipulated, adult males, females and young specimens quickly release a whitish, noxious, sticky and very odorous skin secretion, probably used as defence mechanism against predators (under similar conditions and much longer manipulation times, adult C. asper secrete much less quantities of a much less odorous substance).
Material examined
Type material. HOLOTYPE of C. arnoldi deposited at the Museu de Zoologia de Barcelona ( MZB), Spain, with the following accession number and data: MZB 82-8784 View Materials . Adult male preserved in alcohol; Fogars de Montclús, Montseny, Spain, collected 2 May 1980. Collectors Xavier Pascual and Albert Montori. PARATYPES: MZB 82-8789 View Materials adult male preserved in alcohol; Fogars de Montclús , Montseny collected 2 May 1980. Collectors Xavier Pascual & Albert Montori. MZB 2004-0187 View Materials adult male preserved in alcohol, Fogars de Montclus , Montseny, Abril 1980 ; MZB 2004- 0188 View Materials adult female preserved in alcohol, Fogars del Montclus , Montseny, April 1980 ; MZB 2004-0189 View Materials , adult female preserved in alcohol, Fogars de Montclus , Montseny, April 1980 .
Additional material examined. Apart from the type material preserved in alcohol listed above, morphological observations were based on 59 live specimens from populations A2, A3, B1 and B2. All observations were carried out during the course of an amphibian study from the Natural Park of El Montseny. No additional specimens were preserved for conservation reasons. Instead, data on the morphological appearance of the live specimens was carefully recorded by S.C. and F.A. directly in the field. In total, 20 specimens from locality A2 (10 females 6 males and 4 juveniles), 4 specimens from locality A3 (3 females and 1 male), 32 specimens from population B1 (15 females, 12 males, 3 juveniles and 2 larvae) and 3 specimens from population B2 (2 females and 1 male) where observed and used as a reference for the diagnosis of C. arnoldi (see above) .
Description of the holotype (MZB 82-8784) Measurements: SVL 58.64 mm, HEADL 15.42 mm, DNAR 3.51 mm, DORB 5.02 mm, DES 4.05 mm, DEP 9.32 mm, FLL 15.22 mm, HLL 18.27 mm, AL 24.44 mm, TL 37.56 mm, TH 9.61 mm, TW 3.71 mm.
An adult male. Head strongly flattened, broadest at level of eyes; snout blunt with convex sides in dorsal view, canthus rostralis marked; areas above eyes raised, their width about a third of the distance between them; prominent swellings on the posterior sides of the head present. Body with oval cross-section and some dorsoventral compression; tubercles tipped with hard blunt spines widely distributed on dorsum, becoming smaller and more scattered on head and, on flanks, mainly distributed between the vertical grooves. Cloacal swelling hemispherical with a sagittally elongate opening from which grooves extend laterally. Digits on fore and hind limbs 4:5, not elongate, unwebbed. Tail short (about 64% of snout–vent distance) and strongly compressed laterally, very deep basally (maximum depth 25.5% of length) and tapering gradually to a blunt point.
Colour in alcohol
Dark chocolate brown above with greyish tinge, flanks and sides of tail with scattered irregular, pale grey spots. Underside dark cream, brighter under tail; belly partly translucent with obscure dark markings consisting of dark stippling that are best developed at the sides; throat posterior to gular region with a large dark blotch (immaculate ivory in live specimens, see above). Throat, upper lip margins, underside of tail, and ulnar and palmar aspects of limbs pale and immaculate; tips of digits dark brown.
Other distinguishing features of holotype
A transverse tear across posterior belly extending on to the sides of the body, and a shallow notch on upper border of tail near its tip.
Variation
All living and preserved specimens show the same distinctive traits of the species. The only variation observed is in the pattern of coloration. Some Calotriton arnoldi from populations A1–A3 have irregular pale yellow spots on the tail and body flanks ( Fig. 10B View Figure 10 ), usually more numerous and conspicuous in post-metamorphic newts than in adults (see Fig. 10G–H View Figure 10 ). The extension of pigmentation on the belly is also slightly variable, this being the result of individual variation rather than seasonal variation (our pers. observ.).
Distribution, habitat and conservation
Calotriton arnoldi has only been found in five mountain streams (populations A1–A3, B1 and B2), all within the boundaries of El Montseny Natural Park. Calotriton arnoldi is found between 600 and 1200 m in oligotrophic, cold (below 15 °C) fast running waters preferentially in beech forest ( Fagus sylvatica ) but also in Holm oak forest ( Quercus ilex ), with patches of Algnus glutinosa. In fact, dorsal coloration in all populations of C. arnoldi strikingly resembles that of the submerged leaves of Fagus sylvatica , suggesting it may have a mimetic function.
Although extensive demographic studies have not yet been carried out, preliminary analyses indicate populations A2, B1 and B2 may have very low densities and populations A2 and A3 extremely low densities. Apart from drying out of the mountain streams, other consequences for the low numbers of both specimens and populations observed may be related to reductions in habitable patches as a result of human alteration of its original habitat and replacement of the beech forests by Holm oak forests at medium altitudes (800–1400 m) in response to global warming ( Peñuelas & Boada, 2003). Appropriate conservation measures should be adopted immediately to preserve this interesting Iberian endemic.
Etymology
This new species of newt ( Calotriton arnoldi ) is named after the British herpetologist Dr E. N. Arnold (Department of Zoology, The Natural History Museum, London), for his life-long dedication and contribution to herpetology, and especially to European herpetology. This dedication is also a personal recognition for the 6 years he spent guiding the senior author through his herpetological research while at the Natural History Museum, London.
Proposal of common names in different languages
The following common names for Calotriton arnoldi are suggested:
Catalan: Tritó del Montseny
Spanish: Tritón del Montseny
English: Montseny brook newt


| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Calotriton
| Carranza, Salvador & Amat, Felix 2005 |
Euproctus asper castelmouliensis
| Mertens R & Wermuth H 1960: 17 |
| Mertens R & Muller L 1940: 8 |
Euproctus asper asper
| Mertens R & Wermuth H 1960: 17 |
| Mertens R & Muller L 1928: 9 |
Euproctus asper
| Thorn R 1968: 253 |
| Wolterstoff W & Herre W 1935: 224 |
| Wolterstoff W 1925: 61 |
Euproctus asper var. peyreladensis
| Wolterstoff W 1925: 71 |
Euproctus asper
| Wolterstoff W 1925: 66 |
Euproctus asper var. castelmouliensis
| Mertens R & Wermuth H 1960: 17 |
| Wolterstoff W 1925: 66 |
Euproctus (Hemitriton) asper
| Wolterstoff W 1925: 296 |
Molge bolivari
| Bosca E 1918: 58 |
Triton asper
| Wolterstoff W 1902: 12 |
| Wolterstoff W 1900: 31 |
Molge aspera
| Maluquer J 1916: 58 |
| Boulenger GA 1882: 24 |
Euproctus Pyrenaeus
| Lataste F 1878: 495 |
Calotriton
| Gray JE 1858: 139 |
Calotriton punctulatus
| Gray JE 1858: 139 |
Triton Pyrenaeus Duméril, Bibron & Duméril, 1854: 139
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 139 |
Triton rugosus Duméril, Bibron & Duméril, 1854: 150
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 150 |
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 151 |
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 151 |
Triton puncticulatus Duméril, Bibron & Duméril, 1854: 152
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 152 |
Triton Bibroni
| Dumeril AMC & Bibron G & Dumeril AMC 1854: 153 |
Triton glacialis
| Duges A 1852: 256 |
Hemitriton cinereus Dugès, 1852: 263
| Duges A 1852: 263 |
Hemitriton rugosus Dugès, 1852: 264
| Duges A 1852: 264 |
Hemitriton puncticulatus Duméril 1852
| Duges A 1852: 265 |
Hemitriton punctulatus Dugès, 1852: 265
| Duges A 1852: 265 |
Hemitriton asper Dugès, 1852: 266
| Duges A 1852: 266 |
Hemitriton bibronii Dugès, 1852: 266
| Duges A 1852: 266 |
Euproctus asper ( Dugès, 1852: 1 )
| Garcia-Paris M & Montori A & Herrero P 2004: 120 |
| Montori A & Llorente GA & Santos X & Carretero MA 2002: 48 |
| Salvador A & Garcia-Paris M 2001: 35 |
| Montori A & Carretero MA & Llorente GA & Santos X 1997: 106 |
| Llorente GA & Montori A & Santos S & Carretero MA 1995: 34 |
| Montori A & Campeny R 1991: 1 |
| Montori A 1988: 20 |
| Montori A & Pascual X 1981: 1 |
| Duges A 1852: 1 |