
Austramastodus apterus, Hernando, 2023
|
publication ID |
https://doi.org/10.11646/zootaxa.5315.6.3 |
|
publication LSID |
lsid:zoobank.org:pub:DCD0C570-9808-4198-9998-BBAFD4764982 |
|
DOI |
https://doi.org/10.5281/zenodo.8142490 |
|
persistent identifier |
https://treatment.plazi.org/id/03CCB051-014D-4215-FF6A-C1A247ADFBFC |
|
treatment provided by |
Plazi |
|
scientific name |
Austramastodus apterus |
| status |
sp. nov. |
Austramastodus apterus sp. nov.
( Figs. 1–15 View FIGURES 1–7 View FIGURES 8–15 )
Type locality: Cape Range Peninsula, 22º10’S 113º59’E, Western Australia, Australia GoogleMaps .
Type material: Holotype ♁ ( WAM): “22. 10S 113. 59E WA / N-W Cape Penin. Site / TL-6 18 May–4 June / 1990 J. M. Waldock / CR’90 #84 / pitfall traps ” GoogleMaps . Paratypes: 3 ♀ ( WAM): same data as holotype; 1 ♁ ( NMW): “22. 10S 113.59E WA / N-W Cape Penin. Site / TL-12 3 June 1990 / J. M. Waldock CR’90 / #72 pitfall traps ”; 1 ♁ ( WAM): “22. 12S 113.59E WA / N-W Cape Penin.nr. / Cave, C207 29May / 1990 J. M. Waldock / CR’90 #578” GoogleMaps .
Description. Body. Length: 2.16–2.50 mm (head included), maximum width: 1.16–1.20 mm. Body elongate-oval, dorsal surface covered with dense, golden and silvery pubescence composed of short, and recumbent setae ( Figs. 1–3 View FIGURES 1–7 ).
Head. Punctation granular, very dense, pubescence short and recumbent, distributed radially from the disc ( Fig. 4 View FIGURES 1–7 ); eyes in lateral position, very large and rounded ( Figs. 4–5 View FIGURES 1–7 ), widely separated, space between eyes larger than diameter of one eye ( Fig. 5 View FIGURES 1–7 ); gena with antennal groove ( Fig. 5 View FIGURES 1–7 ).
Thorax. Pronotum transverse, general shape slightly convex; anterior and posterior margins slightly sinuate ( Figs. 1–4 View FIGURES 1–7 ); lateral margins slightly curved; anterior angles broadly acute; posterior angles rounded; disc with coarse and somewhat rough punctation, covered with dense and short, regularly recumbent silvery setae ( Fig. 4 View FIGURES 1–7 ). Elytra long and narrow ( Figs. 1–3 View FIGURES 1–7 ); lateral margins finely bordered along their entire length; apex connected to last sternite by an interlocking device; punctation granular and very dense; pubescence generally silvery, but in some areas with golden spots ( Fig. 1 View FIGURES 1–7 ); setae short and regulary recumbent posteriad; epipleuron broad at base and slightly impressed to receive femoral tips, extending to apical interlocking device. Hind wings absent. Hypomeron broad and slightly depressed, without suture or carina, covered with short silvery setae ( Figs. 2–5 View FIGURES 1–7 ). Prosternal process slightly longer than wide, flat and finely bordered laterally; apex somewhat bluntly pointed. Mesoventrite short and transverse, with large anterior depression to receive prosternal process. Metaventrite uniformly convex, projecting posteriorly, covering the metacoxa. Mesocoxae slightly transverse, trochantins exposed. Metacoxae strongly transverse and oblique, subcontiguous; metacoxal plates large and strongly expanded laterally, posterior margins with two pointed projections, partly covering metatrochantins ( Fig. 2 View FIGURES 1–7 ). Forelegs relatively short and stout, with a few apical spines; tibia clearly shorter than femur ( Fig. 5 View FIGURES 1–7 ). Middle legs relatively short and stout, with a few apical spines; tibia as long as femur. Hind legs very long, with a longitudinal row of many strong spurs; tibia much longer than femur ( Figs. 2–3 View FIGURES 1–7 ). All legs sexually dimorphic, especially protarsus and claws (see below). Tarsal formula 4–4–4.
Abdomen. Ventrites ( Figs. 6–7 View FIGURES 1–7 ) covered with short recumbent setae, finely punctate (not very dense and uniform), and covered by a polygonal squamiform reticulation. Intercoxal process of first abdominal ventrite very small; ventrites 1–3 connate, ventrite 1 long; ventrite 2–4 subequal in length; ventrite 5 with elytral-abdominal interlocking device; setation and apex of ventrite 5 sexually dimorphic (see below).
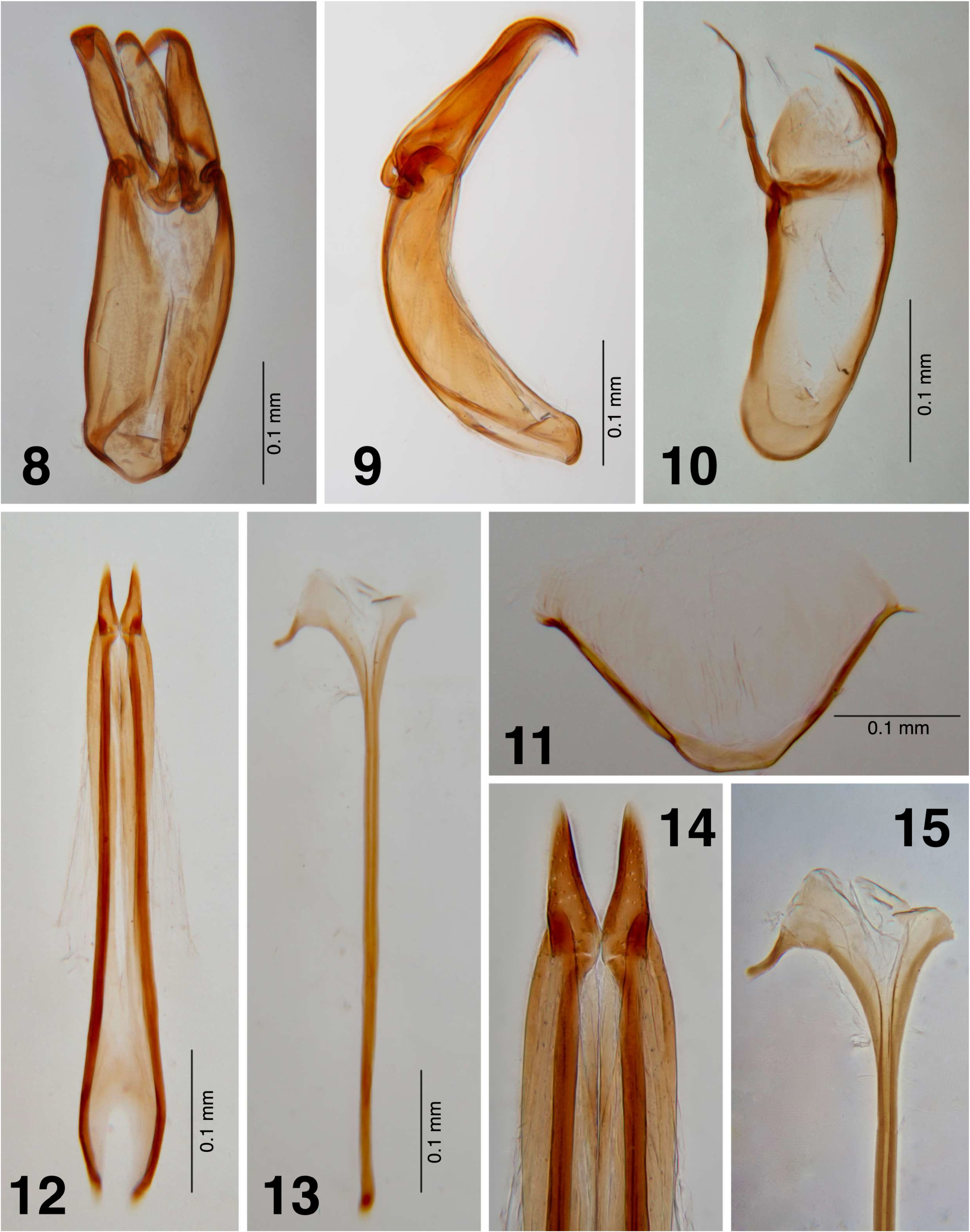
Sexual dimorphism. Male. Ventrite 5 with a pair of sublateral clusters of about 12 long spiniform setae; posterior margin regularly curved apically ( Fig. 6 View FIGURES 1–7 ). Protarsi extraordinarily dilated ( Fig. 5 View FIGURES 1–7 ); claws of the meso and metatarsus asymmetrical. Aedeagus articulated and strongly arched in lateral view ( Fig. 9 View FIGURES 8–15 ), phallobase longer than parameres ( Figs. 8–9 View FIGURES 8–15 ). Median lobe apically strongly curved ventraly, falciform ( Figs. 8–9 View FIGURES 8–15 ). Struts very short, forming a lobe ( Fig. 8 View FIGURES 8–15 ). Parameres slightly longer than median lobe ( Fig. 8 View FIGURES 8–15 ), apex strongly curved ventrad, falciform ( Figs. 8–9 View FIGURES 8–15 ). Genital segment IX spatulate ( Fig. 10 View FIGURES 8–15 ), wide at base; margins strongly sclerotized; apex membranous; parameres of the genital segment IX long, strongly sclerotized and slightly curved, longer than lamina ( Fig. 10 View FIGURES 8–15 ). Sternite VIII arched ( Fig. 11 View FIGURES 8–15 ). Female. Sublateral clusters of ventrite 5 with only one long spiniform seta, remaining setae much shorter ( Fig. 7 View FIGURES 1–7 ); posterior margin acuminate apically ( Fig. 7 View FIGURES 1–7 ). Protarsi not modified, claws of all legs symmetrical. Ovipositor as in Fig. 12 View FIGURES 8–15 , gonocoxal struts very long, ten times as long as gonocoxites, which are strongly acuminate ( Figs. 12–14 View FIGURES 8–15 ); apex of gonocoxal struts articulate. Spiculum ventrale with a long manubrium, distal part with two long lateral expansions ( Figs. 13–15 View FIGURES 8–15 ).
Distribution. So far known only from two very close localities in the central part of the Cape Range Peninsula, Australia ( Western Australia).
Etymology. The epithet, a Latin adjective, means “wingless” and refers to the fact that all specimens collected so far are apterous.
Biology and notes on apterism. According to the data provided by the collector, the holotype and four paratypes were captured with pitfall traps placed in the soil near the entrance of a cave (“C-63”) ( Fig. 16 View FIGURE 16 ), and one specimen was found under stones in the leaf litter of trees (probably Eucalyptus or Corymbia ) near the entrance of another cave (“C-207”, known as “Two Hundred Cave”) (J. M. Waldock, pers. comm.).
The Cape Range Peninsula is an extremely hot and arid region with maximum temperatures of up to 47º C. From 1976 to 1996 the average rainfall was 234 mm ( Environmental Protection Authority, 1999). The two localities are far from the coast (circa 11 km), and there seem to be no permanent aquatic habitats in the surrounding area.
Therefore, we must assume that Austramastodus apterus is the first known terrestrial species of Thaumastodinae .
Perhaps, one of the most relevant peculiarities of the new genus is its apterism, an unusual feature among the largely riparian (paraquatic) Limnichidae usually inhabiting inland water margins and coastal marine habitats. So far, apterism in Limnichidae is known only for a few humicolous species of the subfamily Limnichinae ( Hernando & Ribera, 2003b –c) and for the intertidal and highly specialised genus Hyphalus (Hyphalinae) , which lives on coral rocks ( Hernando & Ribera 2020). Although Doyen (1976) states that all limnichids living in marine habitats are flightless, this is not true for the intertidal Thaumastodinae . The flight ability of some species has been reported from direct observations and can be deduced from the fact that specimens are often collected with light traps ( Spilman 1966; Yoshitomi & Putra 2010; Yoshitomi 2019).
Some hypotheses can be discussed for the absence of wings in Austramastodus apterus . The apterism might, as in other groups of beetles, be an adaptation related to the extremely arid habitat ( Chown et al. 1998). The absence of wings in beetles living in very arid environment is assumed to have evolved in response to reduced environmental heterogeneity or directly to reduce body water loss. Additionally, it could be related to the apparent absence of permanent aquatic habitats in the area, since, as noted above, all other known genera of Thaumastodinae are able to fly and are closely associated with aquatic habitats, whether freshwater habitats, such as wetlands, waterfalls, rivers and streams ( Yoshitomi 2019) or intertidal habitats such as mangroves, coral reefs, dogtooth limestone ( Spilman 1966; Yoshitomi 2019; Liu & Jia 2021), or rockpools at seashores ( Satô 1994). All these habitats have in common a certain instability due to the possibility of sudden water level fluctuations, for instance by unpredictable flash floods in lotic habitats or by storms in coastal areas. Indeed, macroptery appears to be prevalent as a means of escape in certain groups of riparian beetles ( Ramey & Richardson 2017). For example, around 83% of the British terrestrial beetles, which are more or less restricted to wetland habitats, are consistently macropterous ( Lott, 2003), a hypothesis supported by the tendency in some species with wing dimorphism for macropterous specimens to predominate in frequently flooded habitats and brachypterous ones in more stable habitats ( Adis & Junk 2002). Assuming that the newly described species lives in leaf litter or soil and is not associated with any aquatic habitat and therefore not subject to unpredictable instability, might explain its apterism. In any case, further studies will be necessary to determine the habitat requirements of Austramastodus apterus satisfactorily.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Thaumastodinae |
|
Genus |