
Bothriolepis ornata Eichwald, 1840
|
publication ID |
https://doi.org/10.5281/zenodo.4664755 |
|
persistent identifier |
https://treatment.plazi.org/id/03CC6624-FFF8-FFA7-FF66-FE099A68FE6F |
|
treatment provided by |
Felipe |
|
scientific name |
Bothriolepis ornata Eichwald, 1840 |
| status |
|
Bothriolepis ornata Eichwald, 1840 ( Figs 40-54 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
Bothriolepis ornatus Eichwald 1840: 78 .
Bothriolepis prisca – Eichwald 1840: 425.
Bothriolepis ornata – Eichwald 1860: 1513, pl. cvi, fig. 3.
Asterolepis ornata – Eichwald 1861: 448 ( p.p.).
Bothriolepis cf. ornata Eichw. – Gross 1942: 403, 422 ( p.p.) [non text-fig. 11].
For full list of synonyms before 1932, see Gross (1932).
TYPE SPECIMEN. — Woodward (1891) selected MM 116/107 as the lectotype ( Eichwald 1860: 1513, pl. CVI, fig. 3). It is an AMD, collected by Helmersen.
MATERIAL EXAMINED. — From the type locality at Priksha River: MM 5/198, PVL ; BMNH P.4600, ML1, P.710, MxL; PIN 835 View Materials /4, Nu. From the locality at Skujaine River near Klūnas hamlet: LDM 43 View Materials /730, 100/1-65, 145-147, 336, 346, 354-368, 396-426, 441, 523, 525-527, LGI 5 /2045-2078, two articulated headshields and their fragments, disarticulated plates of the head-shield, trunk-armour and pectoral appendage ; PIN 1491 View Materials /89, ADL, 1491/90-95, two Pn, two Sm, Pmg, CD 2 .
LOCALITIES AND HORIZON. — The type locality is the exposure at Priksha River, Russia; the upper Famennian Lnyanka Beds. Other studied material comes from the two outcrops of white and pink sand containing abundant fish bones, dolomite marl and marl at the right bank of Skujaine River down Klūnas village, Latvia; the Famennian Tērvete Formation. Studied fragments of plates, undescribed here, which comes from Msta River near Beryozovik village (kept at IEC), and those reported from Belaya, Lnyanka and Mshanka Rivers in Novgorod region ( Ostrometskaya & Kotlukova 1966; Obruchev 1964) and the North Timan, the Pokayama Formation ( Obruchev 1958), probably also belong to B. ornata .
DIAGNOSIS. — Rather large Bothriolepis with the dorsal length of the trunk-armour reaching 230-240 mm and the length of the head-shield at least 100 mm. B/L index of head-shield 127. Preorbital recess of trifid type. Orbital edges of Prm and La are thickened. Prm of moderate breadth, with B/L index about 112, rostral margin 1.7-1.9 times longer than orbital margin. Nu arched, narrow, with L/B index of 83. Pn narrow. B/L index of dorsal wall of trunk-armour about 82. Median dorsal ridge poorly developed. AMD moderately narrow, B/L index about 79-88. Anterior margin of AMD short and concave. PMD narrow, B/L index about 75-89, 82 on average. ADL with narrow lateral lamina and large postnuchal ornamented corner. Axillary foramen long and narrow. Proximal segment of the pectoral appendage moderately long, four times as long as broad. Ornamentation typically reticulate, in quite large individuals becoming coarser and sparser.
DESCRIPTION
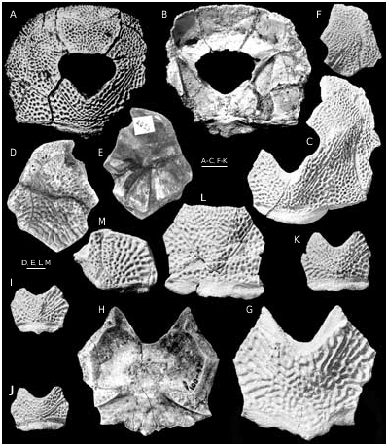
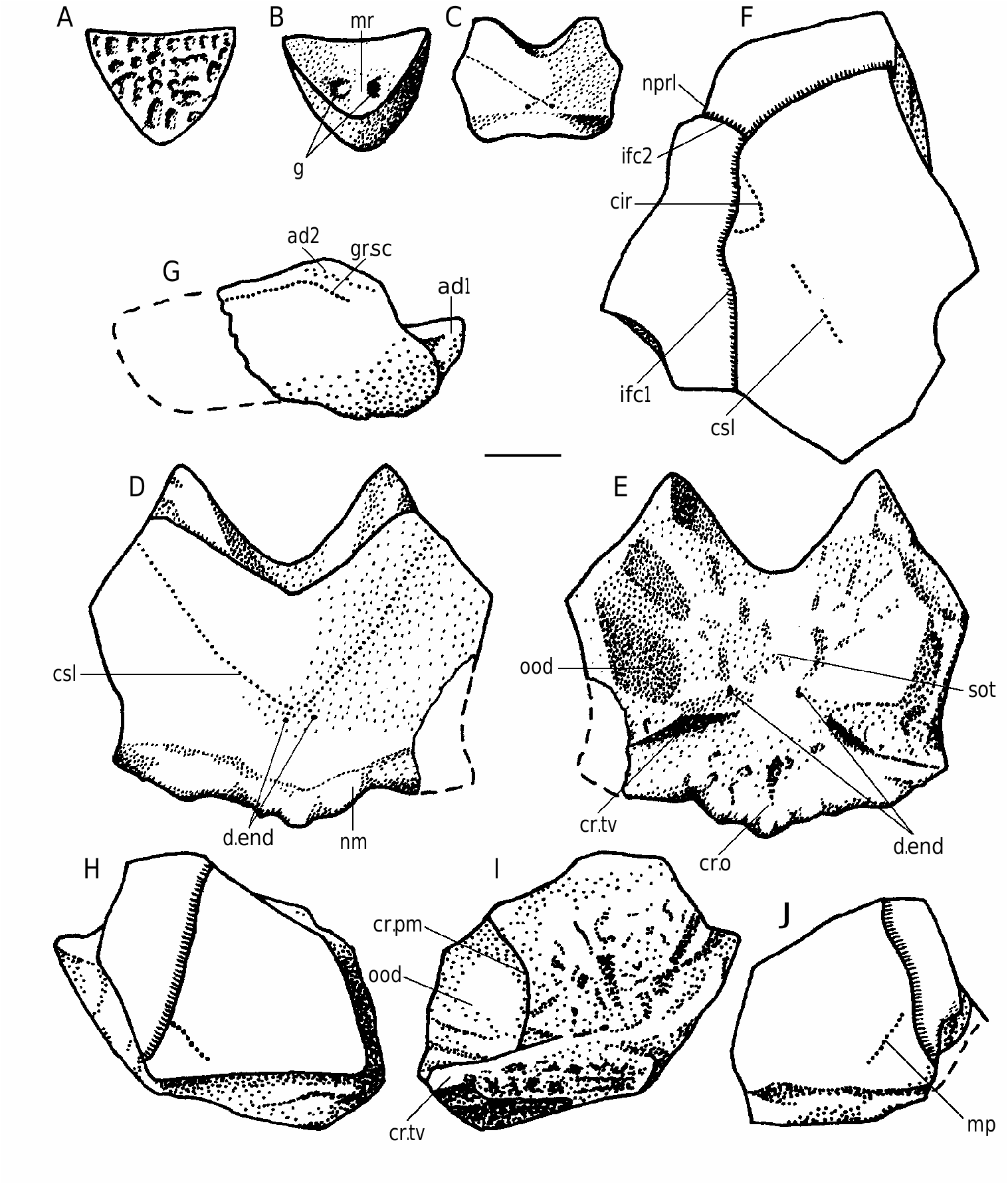
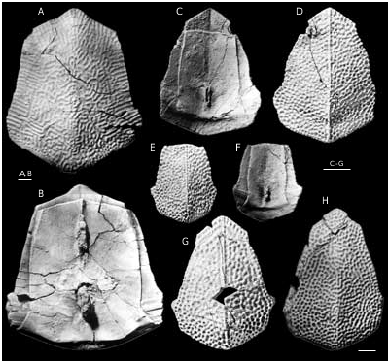
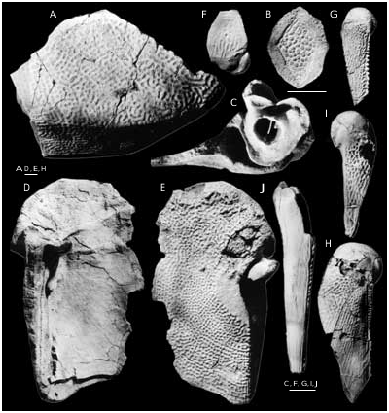
Bothriolepis ornata is well-represented at the locality at the right bank of Skujaine River 1 km E from Klūnas hamlet by many disarticulated plates and some articulated skulls. Most are from well-grown individuals of moderate size, but there are also some small, and quite large specimens. Bothriolepis ornata is one of the largest of species of Bothriolepis , but did not quite reach the size of B. maxima or B. gigantea . The head-shield ( Figs 40 View FIG A-C; 42A, B) has a B/L index of about 127, and is strongly vaulted both rostrocaudally and transversely. The rostral margin is convex, slightly shorter than the posterior margin, which is weakly convex and bears a welldefined posterior median process. There are well-defined anterolateral corners (alc) and a deep prelateral notch (nprl). The obtected nuchal area (nm) is long, extending onto the Pn, it is broadest on the Nu. The orbital fenestra is relatively short and broad (B/L index about 200). Preorbital recess (prh) is distinctly trifid with extended lateral horns and a point- ed median division, as in B. maxima and B. hayi Miles, 1968 . The median division reached the middle of Prm, but lateral horns did not reach the middle of La, as is the case in B. hayi . The orbital edges of the Prm and La are thickened, as in B. ciecere , B. macphersoni and B. karawaka .
The visceral skull surface ( Fig. 40B View FIG ) shows the broad otico-occipital depression which is welldefined by the paramarginal cristae. The anterolateral corner of otico-occipital depression is broad in its base and extended nearly to the rostral margin of the orbital fenestra. The posterolateral corner is rounded and does not extend laterally over the middle of the Pn’s posterior margin, as it does in B. ciecere . The transverse lateral groove is moderately broad and clearly defined. A relatively broad shallow depression anteriorly from the antero-lateral corner of preorbital recess is the lateral pit, which is situated more laterally than mesially. The median occipital crista is relatively low, often it consists from several small ridges. The transverse nuchal crista is prominent. The median ridge (mr) sharing the broad paired pits (g) of Pp plate is broad and anteriorly bears a tubercle. The supraotic thickening on the Nu is very low.
The Prm ( Fig. 42 View FIG C-E) is broadest slightly posteriorly from the infraorbital sensory groove. The rostral margin is convex, the orbital margin is straight or weakly concave and bears clearly defined but shallow nasal notch (pnn). The infraorbital sensory groove (ifc1) crosses the plate anteriorly from its middle part. The shape and proportions of the Prm resemble that in B. ciecere , differing well from that in such taxa as B. leptocheira and B. maxima .
The La ( Figs 40 View FIG D-F; 41; 43F) is moderately broad with L/B index about 123-139, 132 on the average. The rostral margin is relatively short and almost straight, the antero-median and antero-lateral corners are well-defined. The infraorbital sensory groove (ifc1) crosses the plate in its anterior part not far from the lateral margin. The central sensory line groove (csl) usually finishes slightly anteriorly the middle of an orbital fenestra length, it might be interrupt- ed or very short.
The Pp ( Figs 40L View FIG ; 43A, B View FIG ) is broad, L/B index varies from 56 to 83. As in other species the anterior margin is strongly convex in small specimens and became almost straight with increasing size.
The Nu ( Figs 40 View FIG G-L; 43C-E) is vaulted with an angle between right and left halves about 132°. It is relatively narrow, L/B index 72-93, 83 on the average, and in most aspects resembles that in B. ciecere . The anterior division of the lateral margin usually is concave and equal or a little longer than the posterior division. The posterior margin is weakly convex and bears well-defined median process (prnm). The orbital facetes are short, similarly as in B. leptocheira and B. ciecere . There are short supraoccipital grooves (socc), which terminate little in front of the obtected nuchal area at the external openings for the endolymphatic ducts (d.end). Specimen LDM 100/42 ( Figs 40G View FIG ; 43D View FIG ) shows the broad unornamented area along the postpineal notch, which probably was overlapped by the extremely broad Pp. As a result the outer surface of this Nu was excluded from contact with the orbital fenestra, as in Asterolepis .
The Pn ( Figs 40M View FIG ; 43 View FIG H-J) with L/B index about 98. The lateral division of the Pn is relatively narrow and composing 50% of the breadth of a median division of the plate. The median division is as long as it is broad. The lateral margin of the plate is short. The Pn usually bears a short middle pit-line groove (mp).
The Sm (extralateral plate) is known from two badly preserved specimens PIN 1491/92 ( Fig. 43G View FIG ), and PIN 1491/93, both are the anterior end of the plate. The dorsal margin has a prominent antero-dorsal process (ad1). There is a groove (gr.sc) running along the dorsal margin of the ornamented part of the plate, which is similar to that of B. obrutschewi (see KaratajūteTalimaa 1966) and B. macphersoni .
The trunk-armour description is based on plasticine reconstruction composed of large disarticulated plates. The trunk-armour is relatively low and broad (B/L index 82), and broader than in B. leptocheira or B. ciecere (B/L index in both species about 77). Length of the dorsal wall probably exceeded 240 mm in largest individuals, but was only 50 mm in the smallest examined specimens. The dorsal wall is of moderate height, with right and left laminae enclosing an angle of about 132°. The median dorsal ridge is weakly defined, the dorsal wall is rounded transversely. The dorsolateral and ventro-lateral ridges are welldefined, but in quite large individuals slightly rounded. The characteristic feature is that the depression anteriorly the tergal angle is weakly developed as in B. groenlandica Heintz, 1930 (see Stensiö 1948).
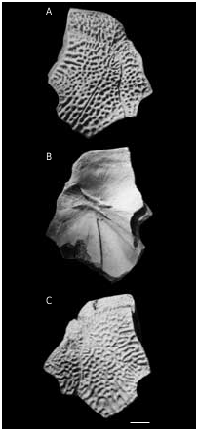
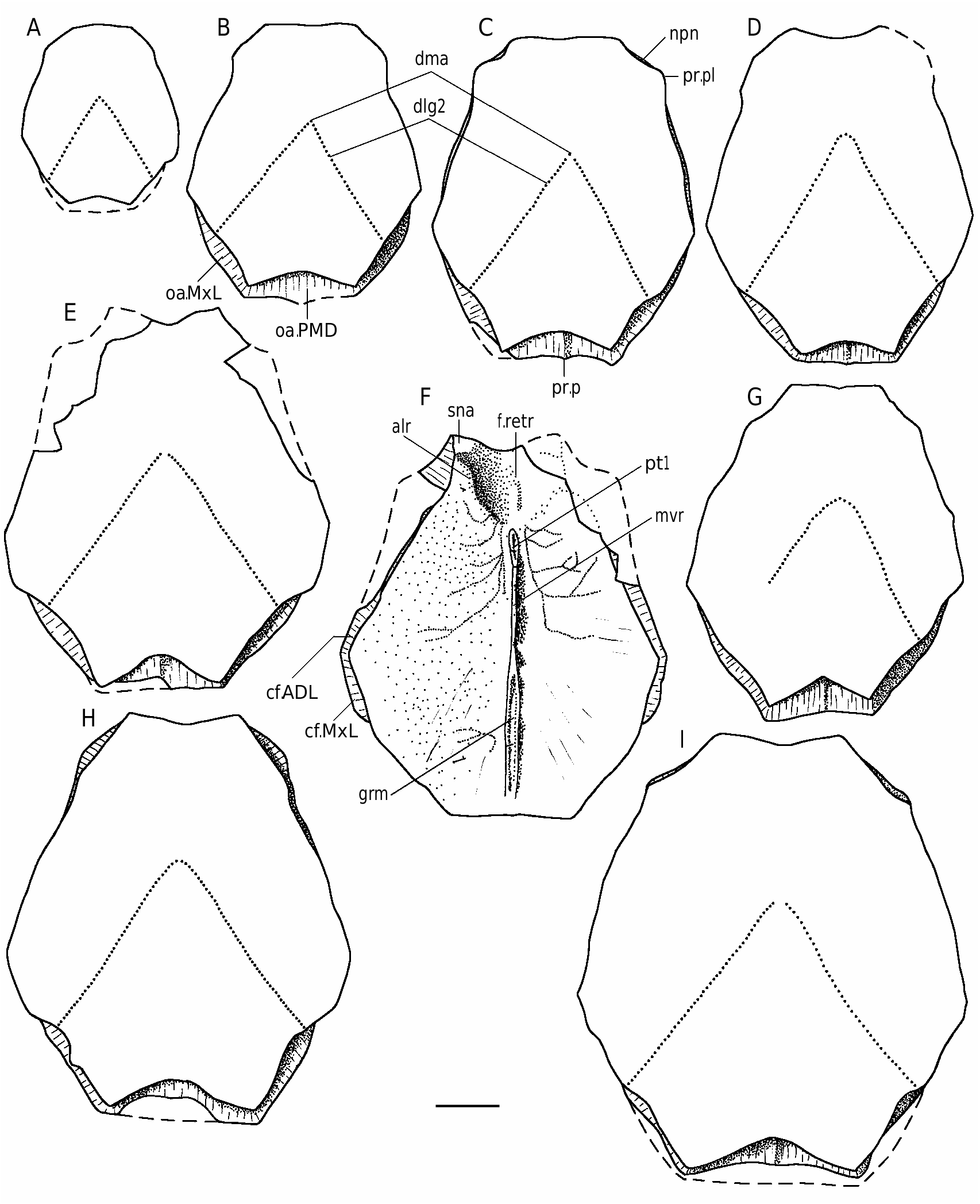
The AMD ( Figs 44-46 View FIG View FIG View FIG ) with B/L index about 79-88, 83 on the average. The anterior margin is always concave. It is narrow, usually shorter than the posterior margin, and 2.2-3.1 times narrower as a total breadth of the plate, differing in its proportions from all the other species of Bothriolepis from the Famennian of Baltic area. The antero-lateral and lateral corners are round- ed, the postnuchal notch (npn) is deep and postlevator process (pr.pl) is sharply defined. The posterior division of the lateral margin is 1.4-1.6 time shorter than the anterior division. There is no median dorsal ridge. Overlap areas for ADL and MxL are normally developed as usually in Bothriolepis in general, but in large individuals the sutural connection of AMD with MxL is often of Remigolepis type. The overlap area for ADL along the postnuchal notch is of wedge type: the outer surface of ADL slightly overlaps AMD. The anterior oblique dorsal sensory line groove (dlg1) is not present even in smallest specimen, the posterior oblique dorsal sensory line groove (dlg2) is well-defined also on the plates of rather large individuals. In some cases the posterior oblique dorsal sensory line groove is short: specimen LGI 5/2078 ( Fig. 46H View FIG ) shows dlg2 on the left side terminat- ed in front of the postero-lateral margin.
The visceral surface of the AMD ( Figs 44B View FIG ; 45B, E View FIG ; 46F View FIG ) shows a narrow elongated levator fossa (f.retr), which is delineated by the low and narrow postlevator thickenings (alr). The supranuchal area (sna) is well-defined and broadest at the antero-lateral corners. The anterior ventral pit (pt1) is deep. The median ventral ridge (mvr) is high, it divides in the posterior third of the plate to form a deep median ventral groove (grm).
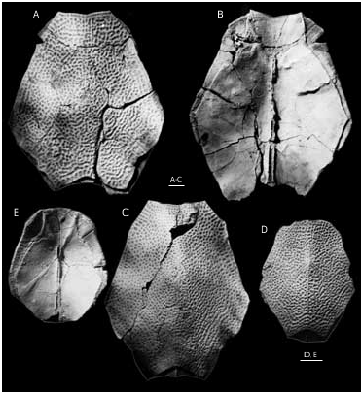
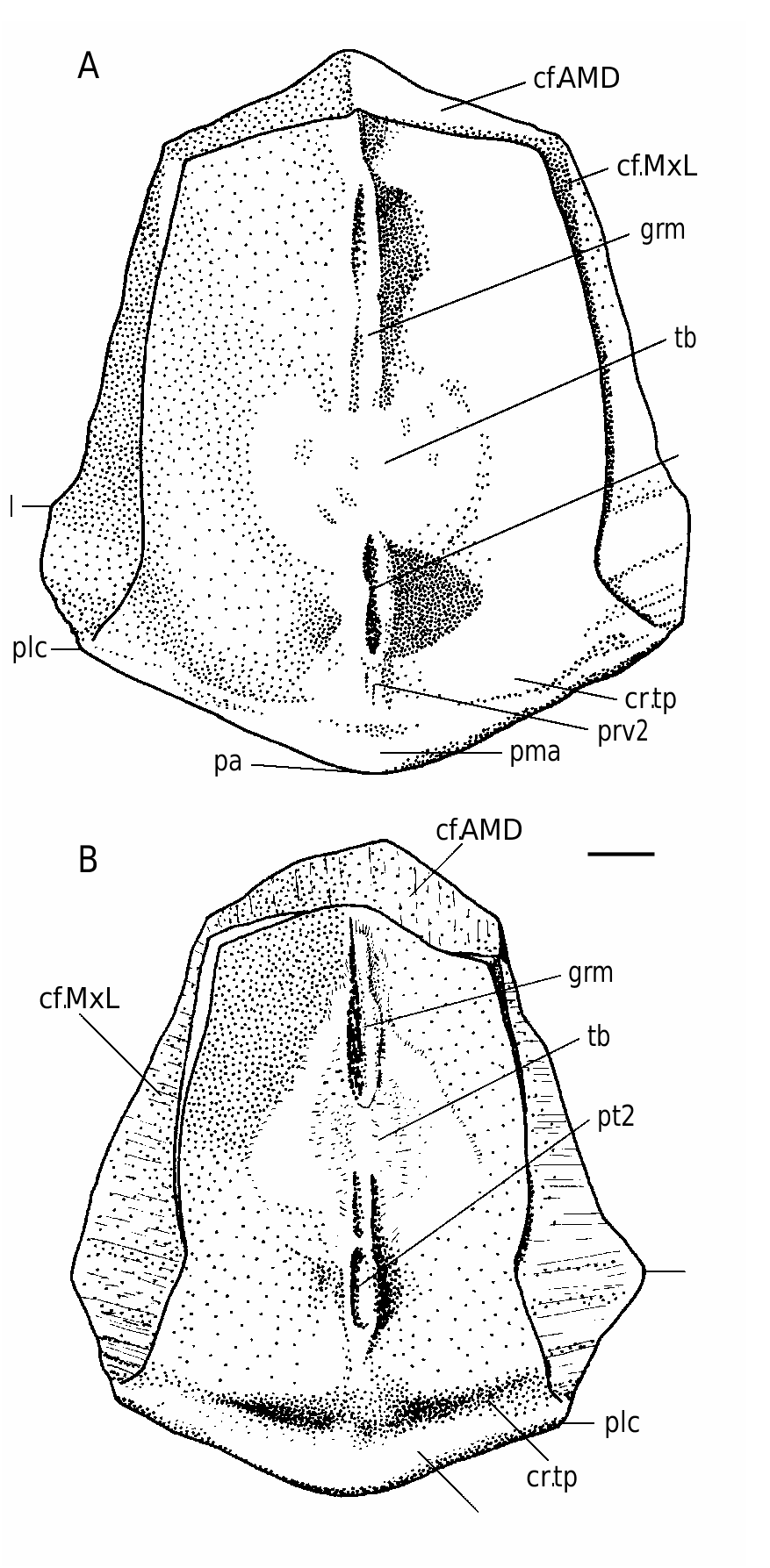
The anterior and posterior margins of the PDM ( Figs 47-49A, B View FIG View FIG View FIG ) both are strongly convex, with a well-developed anterior and posterior corners. The lateral and postero-lateral corners are also well-defined. The width of the anterior margin varies between 45-63% of total breadth of the plate. The PMD is arched with the median dorsal ridge (dmr) well-defined in individuals of small size; in large individuals the dmr is rounded. The suture with the MxL is of wedge type. The median ventral ridge and median ventral groove are weakly defined on the visceral surface of the plate in small individuals and the median ventral ridge is rather high with the deep posterior ventral pit (pt2) and strongly developed ventral tuberosity (tb) in large individuals ( Fig. 48 View FIG ). The crista transversalis interna posterior (cr.tp) is low and smoothed, the postmarginal area (pma) being narrow.
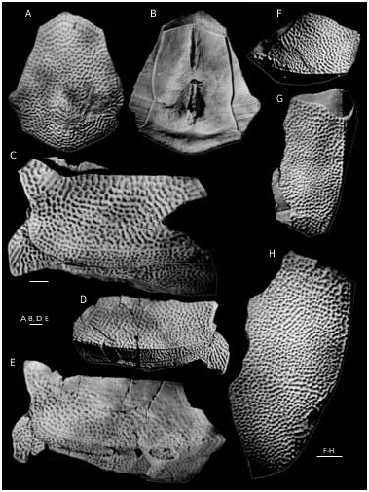
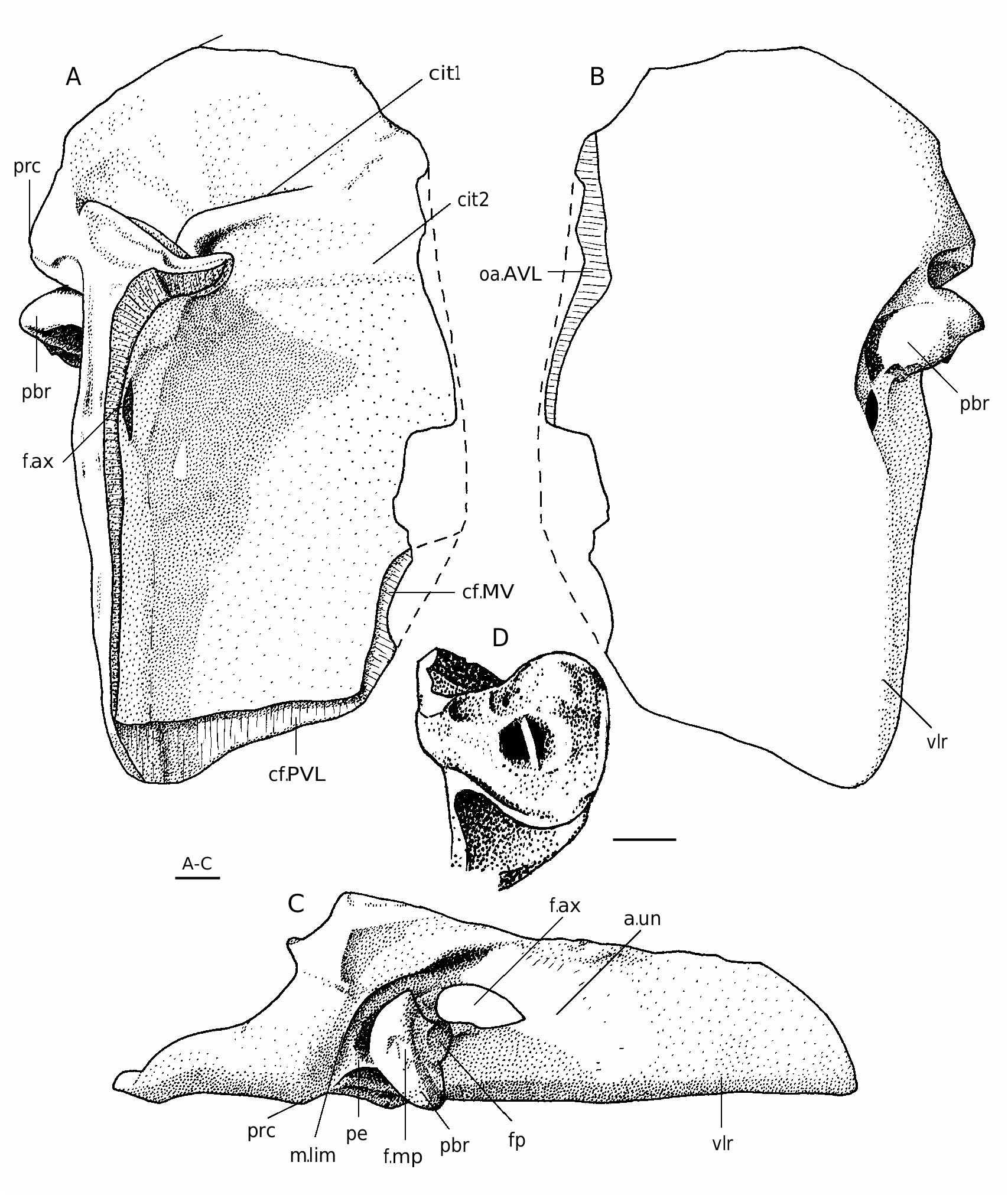
The dorsal lamina (dlm) of the ADL ( Figs 49 View FIG C-E; 50A-C) is relatively long, but due to significant breadth of a strongly pronounced postnuchal ornamented corner (pnoa) it is only 2.2 as long as it is broad. The lateral lamina is three times as long as it is high. The ADL is not simply overlapped by AVL, but also at the same time overlaps that plate posteriorly the processus obstans, as in B. canadensis , B. groenlandica and B. maxima .
The MxL ( Figs 49F View FIG ; 50 View FIG D-F; 52A) is moderately broad. The dorsal lamina of the plate is less than twice (1.7-1.8 time) as long as it is broad. The lateral lamina is 2.5-2.6 times as long as it is broad, with long and sharp postero-ventral ornamented corner (cu).
The AVL ( Figs 51 View FIG A-C; 52D-E) is of moderate breadth, the ventral lamina of the single complete plate is 1.5 time as long as it is broad. The subcephalic division is of moderate length, comprises 29% of total length of the ventral lamina and has a weakly defined antero-lateral corner (c.al). The right AVL overlaps the left similar to the other Bothriolepis . The axillary foramen (f.ax) is rather large and about twice as long as it is broad ( Fig. 51C View FIG ). The processus brachialis shows some features not found in other Bothriolepis from the Main Devonian Field ( Fig. 51D View FIG ). The fossa articularis pectoralis is not boynded off anteriorly by the margo limitans. The groove around the external opening of the funnel pit (fp) is deep, its dorsal part is divided into two unequal deep pits; the larger and more distally situated pit bears an opening of the canal. Specimen LDM 100/365 ( Figs 51D View FIG ; 52E View FIG ) shows the funnel pit divided into two divisions by very thin longitudinal wall. Probably this structure is the result of the joint desease.
The visceral surface of the AVL ( Figs 51A View FIG ; 52D View FIG ) shows as in most part of other species of Bothriolepis the high transverse anterior crista (cit1) running antero-mesially and the low and broad transverse thickening (cit2) directed more mesially.
The PVL ( Fig. 49G, H View FIG ) is of moderate breadth, the ventral lamina is twice as long as it is broad. The subanal division is relatively narrow, it occupies about 30% of the total PVL length. The lateral lamina is about twice as long as it is high. Left PVL overlaps the opposite PVL.
The MV ( Fig. 52B View FIG ) is more elongated than in most species of Bothriolepis , the L/B index reaching about 1.4-1.7.
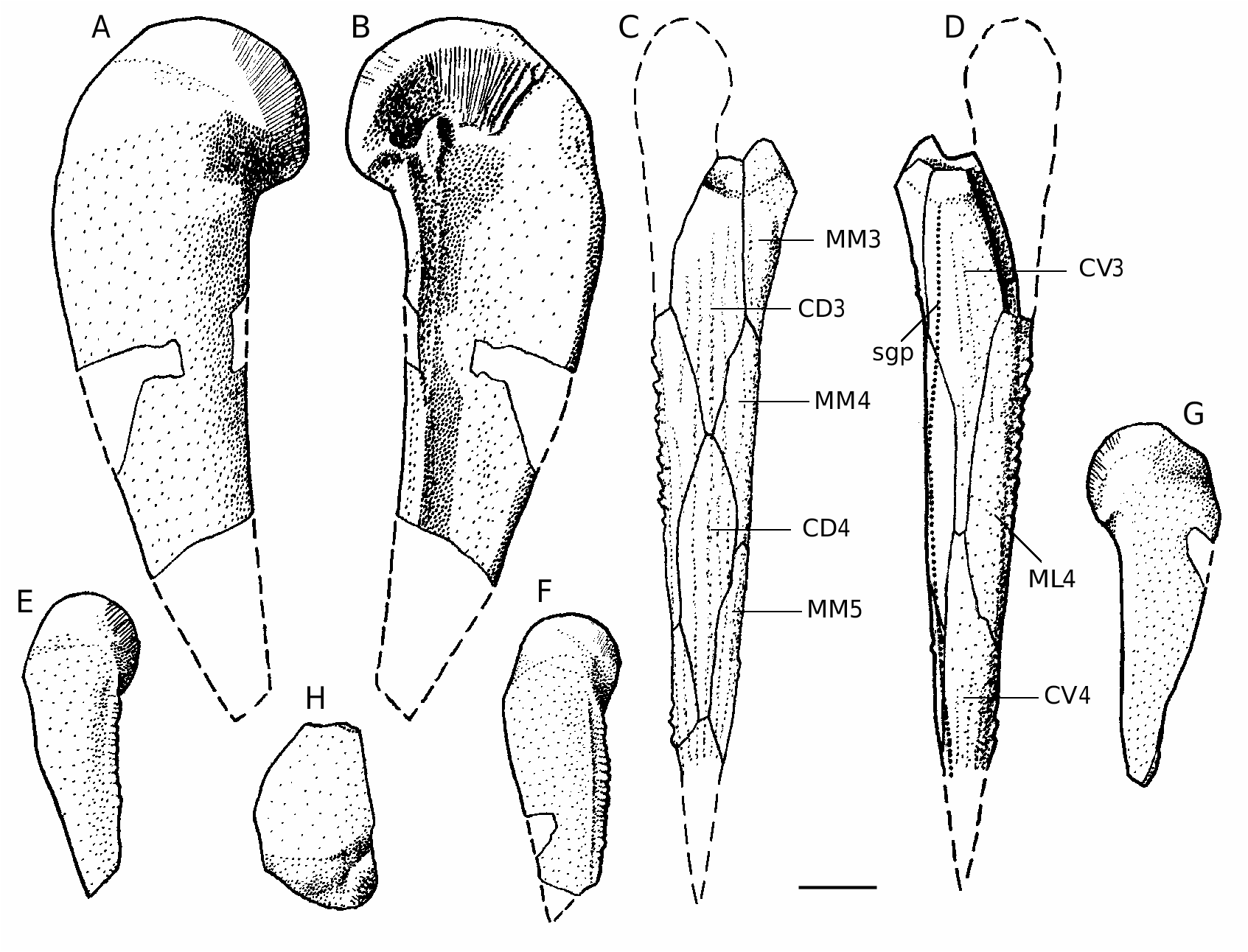
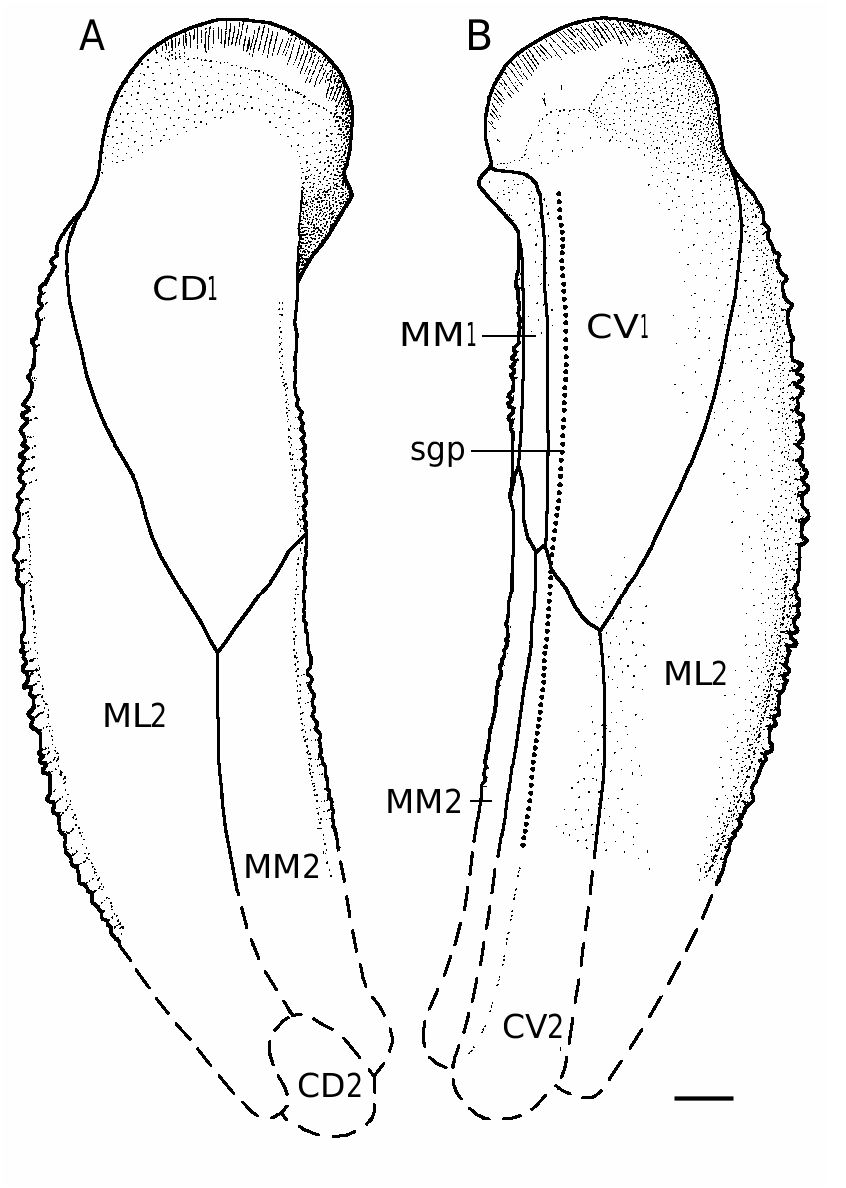
The pectoral fin is represented by many disarticulated bones ( Figs 52 View FIG F-I; 53), and two specimens showing articulated plates of the proximal segment associated with fragments of AVL ( Fig. 54 View FIG ). There are three examples of the distal segments ( Figs 52J View FIG ; 53C, D View FIG ). Both segments bear prominent lateral and mesial spines. The spines are large and closely setting on the proximal segment, in large individuals they are fusing at the base. The ventro-mesial margin is smoothed and rounded. The pectoral pit-line groove (sgp) is traced almost along the ventro-mesial margin and can be seen not on the MM2 as in B. canadensis , but on the CV1 and CV2 similar to B. cristata Traquair, 1895 and B. ciecere . The lateral spines of the distal segment are sharp and proximally directed, the mesial margin bears few rounded tubercles. The CD1 is moderately broad with L/B index about 2.7. The CV1 is slightly more elongate than the CD1 (L/B 3.1). The CD2 is slightly longer than broad. The CV2 is 4.5-5.1 times as long as it is broad. The distal segment is normally developed, showing the CD3 and CD4, but CD5, which is well-defined in B. leptocheira , in B. ornata is not present.
The ornamentation is typically reticulate, in quite large individuals becoming coarser and sparser. The network of anastomosing ridges in large individuals is broken into irregular ridges, but never into short ridges or tubercles. The irregular ridges are situated without any order, only one specimen of PMD shows ridges perpendicular to margins of the plate. The ornamentation of the pectoral appendage remains reticulate even in quite lagre individuals. The distal segment ornament consists of low longitudinal ridges, which are weakly defined on the ventral surface and more distinct on the dorsal surface of the segment.
REMARKS
Bothriolepis ornata was introduced by Eichwald (1840) for specimen of the AMD collected by Helmersen at Priksha River (as indicated on the label), tributary of Msta River, near Borovichi town in the Novgorod region. The lectotype and four other specimen from the type locality (collected by H. Helmersen, A. Olivieri & R. Hecker) were studied in details and compared with the material from Skujaine River, Latvia, collected by the author (hold in LDM) and Karatajūte-Talimaa (kept in LGI and PIN), and attributed to the same species by Lukševičs (1992). The following account is the first full treatment in English, all materials from the type locality and Latvia are described here.
DISCUSSION
Bothriolepis ornata at the Skujaine locality is associated with B. jani (Lukševičs 1986) . B. ornata can be distinguished readily from B. jani by its 1) larger size; 2) typical reticulate ornament, which is tubercular in B. jani ; 3) shape and proportions of almost all trunkarmour plates. B. ornata also resembles B. ciecere by some features (see description of B. ciecere for comparisons).
Bothriolepis hayi is a species that most resembles B. ornata among Scottish Bothriolepis . B. ornata differs from B. hayi most strikingly in the larger size and the narrower AMD and PMD, but also in the proportions of the Prm, La, Nu, Pn and more broad postnuchal ornamented corner of the ADL.
B. ornata is morphologically close to B. groenlandica ( Stensiö 1948) . The two species are of similar size, and have similar proportions and shape of almost all the head-shield, trunk-armour and pectoral appendage plates. The distinctions between them are insignificant and consist in that in B. ornata 1) the head-shield is slightly narrower and more arched; 2) the La and Pn are narrower; 3) the rostral margin of the La is shorter; 4) the mesial division of the Pn is of different shape. These could be intraspecific variations or preservation differences, but direct comparison of the Baltic and Greenland material is needed to solve this problem.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bothriolepis ornata Eichwald, 1840
| Lukševičs, Erwin 2001 |
Bothriolepis cf. ornata Eichw.
| GROSS W. 1942: 403 |
Asterolepis ornata
| EICHWALD E. 1861: 448 |
Bothriolepis ornata
| EICHWALD E. 1860: 1513 |
Bothriolepis ornatus
| EICHWALD E. 1840: 78 |
Bothriolepis prisca
| EICHWALD E. 1840: 425 |