
Sattleria karsholti, Huemer, Peter & Hebert, Paul D. N., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.278322 |
|
DOI |
https://doi.org/10.5281/zenodo.5630997 |
|
persistent identifier |
https://treatment.plazi.org/id/03CA8790-FFBC-1669-FF4F-FE3FEF2C0D92 |
|
treatment provided by |
Plazi |
|
scientific name |
Sattleria karsholti |
| status |
sp. nov. |
Sattleria karsholti View in CoL sp. nov.
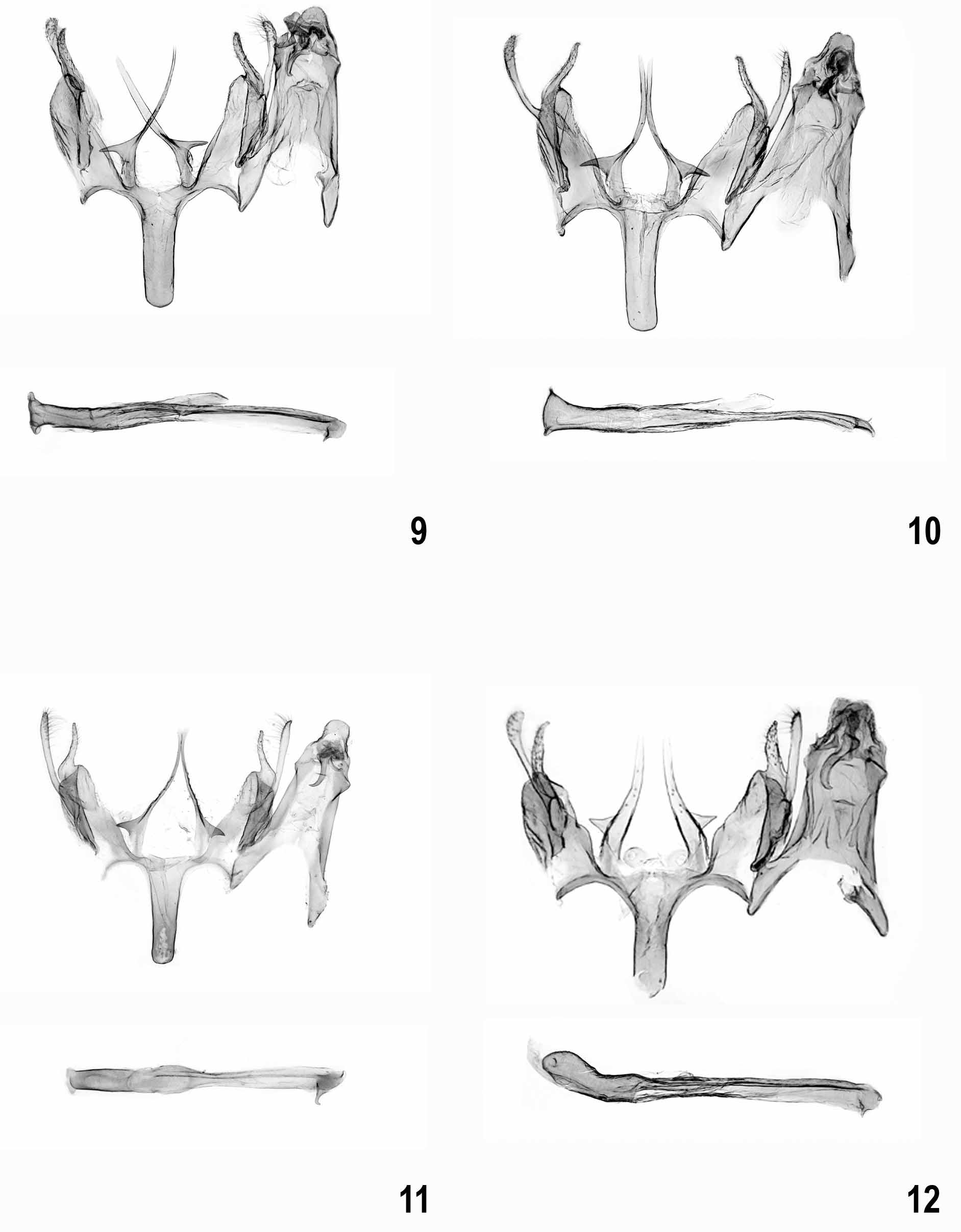
( Figs 3 View FIGURES 1 – 8 , 12 View FIGURES 9 – 12 , 24–25 View FIGURES 24 – 27 )
Type material. Holotype 3, ‘ Italia sept. Prov. Bergamo Alpi Orobie Val d´Arera 2100 m 14.- 15.8.1992 leg. Huemer’ ‘ BC TLMF Lep 1439’ ( TLMF).
Paratypes. Italy: 1 3, same data as holotype, gen. slide GEL 1132 ( TLMF); 7 3, Prov. Trento, Adamello, Mandron, 2800 m, 30.7.- 1.8.1964, leg. Burmann, gen. slides GEL 131, LMP 52, LMP 71 ( LNK; TLMF; ZMUC); 2 3, same data, but end 7.1967, leg. Burmann; 1 3, same data, but 2700 m, mid 8.1958, leg. Burmann, gen. slide LMP 89 ( LNK); 1 3, same data, but 2500 m, 15.8.1985, leg. Schütz ( RCTG); 1 3, same data, but 2600 m, 15.8.1989, leg. Schütz ( RCTG); 1 3, Prov. Verona, Monte Baldo, Telegrafo, 2150 m, mid 7.1969, leg. Burmann, gen. slide LMP 74 ( LNK); 1 3, Prov. Verona, Monte Baldo, Cima Valdritta, 2200 m, 15.7.1987, leg. Huemer & Tarmann, gen. slide GEL 162 ( TLMF); 1 3, Prov. Verona, Monte Baldo, Longino, 2200 m, 29.6.1985, leg. Tarmann ( TLMF).
Description. Adult ( Fig. 3 View FIGURES 1 – 8 ). Head cream-coloured, rarely mid-brown, labial palpus cream-coloured, with few mid-brown scales on outer surface, tip of segment three brownish; antenna blackish brown; thorax and abdomen mid-brown, mixed with some rusty brown. Wingspan 3 17.5–20.5.0 mm; forewing ground colour underlies some variation, from light creamy brown to darker grey-brown, rusty brown along subcosta, fold and in basal half of forewing, medial part of wing intensively mottled cream, indistinct angulate cream fascia at 4/5 inwardly bordered by irregular transverse dark brown fascia; black markings: dash in fold, subcostal spot at 2/5 and angulated spot at 3/ 5 in middle of forewing, furthermore some black mottling at base and along costa, termen with black dots; fringes concolorous with ground colour, weakly defined fringe line present; hindwing light grey with concolorous fringes. Female unknown.
Male genitalia ( Figs 12 View FIGURES 9 – 12 , 24–25 View FIGURES 24 – 27 ). Uncus with evenly rounded apex; gnathos hook strong, culcitula large; tegumen anteriorly widened, broadly and deeply emarginated anterior margin; pedunculi long, slender; valva long, slender, extending almost to apex of uncus, nearly straight; sacculus shorter than valva, evenly tapered to apical point, basally without lobe; vinculum deeply emarginated with pair of long processes; primary process long and broadly digitate, distal half with few fine setae, apically weakly narrowing, almost level with apex of sacculus; secondary process short, sub-triangular spine, arising at right angle in basal third of primary process; saccus slightly shorter than primary vincular process, slender sub-rectangular with cut-off apex; anellus with pair of small, rounded, centrally perforated sclerites; phallus slender, nearly straight, with small medial projection, coecum weakly inflated, apex with short and straight sclerotized arm.
Female genitalia. Unknown.
Diagnosis. Sattleria karsholti sp. nov. is externally very similar to other medium-sized species of the genus, particularly to the south-western alpine population of S. melaleucella . However, S. melaleucella has a longer and more slender primary process of the vinculum, a longer spine-like secondary process and a large medial projection of the phallus ( Figs 9–10 View FIGURES 9 – 12 , 26–27 View FIGURES 24 – 27 ). In the somewhat similar S. arcuata , the primary vincular process is needleshaped and serrated ( Fig. 11 View FIGURES 9 – 12 ). Intraspecific divergence at COI is absent in the two specimens examined whereas interspecific divergence is 2.34% to the nearest neighbour S. marguareisi . The minimum distance to the allegedly conspecific S. melaleucella is higher at 4.11%.
Bionomics. Host-plants and early stages are unknown. The adults have been collected from the end of June to mid-August. Habitats are alpine scree and rock formations primarily on limestone, but the species also occurs on siliceous soil with sparse vegetation at elevations ranging from about 2200 m to 2800 m.
Distribution. Only known from a small section of the southern Alps, ranging from Monte Baldo in the west to Pizo Arera in the east (Prov. Trento, Verona, Bergamo, Italy).
Etymology. The species is named after our colleague and friend Ole Karsholt (Zoological Museum, Copenhagen) in recognition of his outstanding contribution to European lepidopterology.
Remarks. S. karsholti sp. nov. was hitherto considered as the geographical form “C” of S. melaleucella , representing this species in the southern Alps ( Pitkin and Sattler, 1991; see discussion). Despite the presence of several distinctive morphological characters, Pitkin and Sattler (1991) hesitated to introduce a new name in the absence of further evidence indicating species status.
Pl. 21, Fig. 207c in Huemer and Karsholt (2010) depicts S karsholti sp. nov. and not S. melaleucella .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |