
Sattleria Povolný, 1965
|
publication ID |
https://doi.org/ 10.5281/zenodo.278322 |
|
DOI |
https://doi.org/10.5281/zenodo.5630992 |
|
persistent identifier |
https://treatment.plazi.org/id/03CA8790-FFBB-166A-FF4F-F9BAEB590B8D |
|
treatment provided by |
Plazi |
|
scientific name |
Sattleria Povolný, 1965 |
| status |
|
Genus Sattleria Povolný, 1965 View in CoL
Type species: Gelechia dzieduszyckii Nowicki, 1864 by monotypy and original designation.
Diagnosis. Adult ( Figs 1–8 View FIGURES 1 – 8 ). Head smoothly scaled, labial palpus recurved, second segment with short brush of scales ventrally, third segment acute, about length of second segment. Wingspan 3 15–24 mm, Ƥ 10–19 mm. Male with longish forewing, maximum width at about 4/5; hindwing broad. Female with short and distinctly pointed forewing, maximum width in basal third; hindwing largely reduced, short and pointed flap of about 1/3 length of forewing. Forewings of both sexes light grey brown to mid-brown, frequently with darker mottling in basal third and at about 2/3; dark brown markings consisting of basal streak or one to two elongate black stigmata in fold and further spots at middle and end of cell, the latter frequently angulated; termen often with black dots, particularly in male. Hindwings light grey.
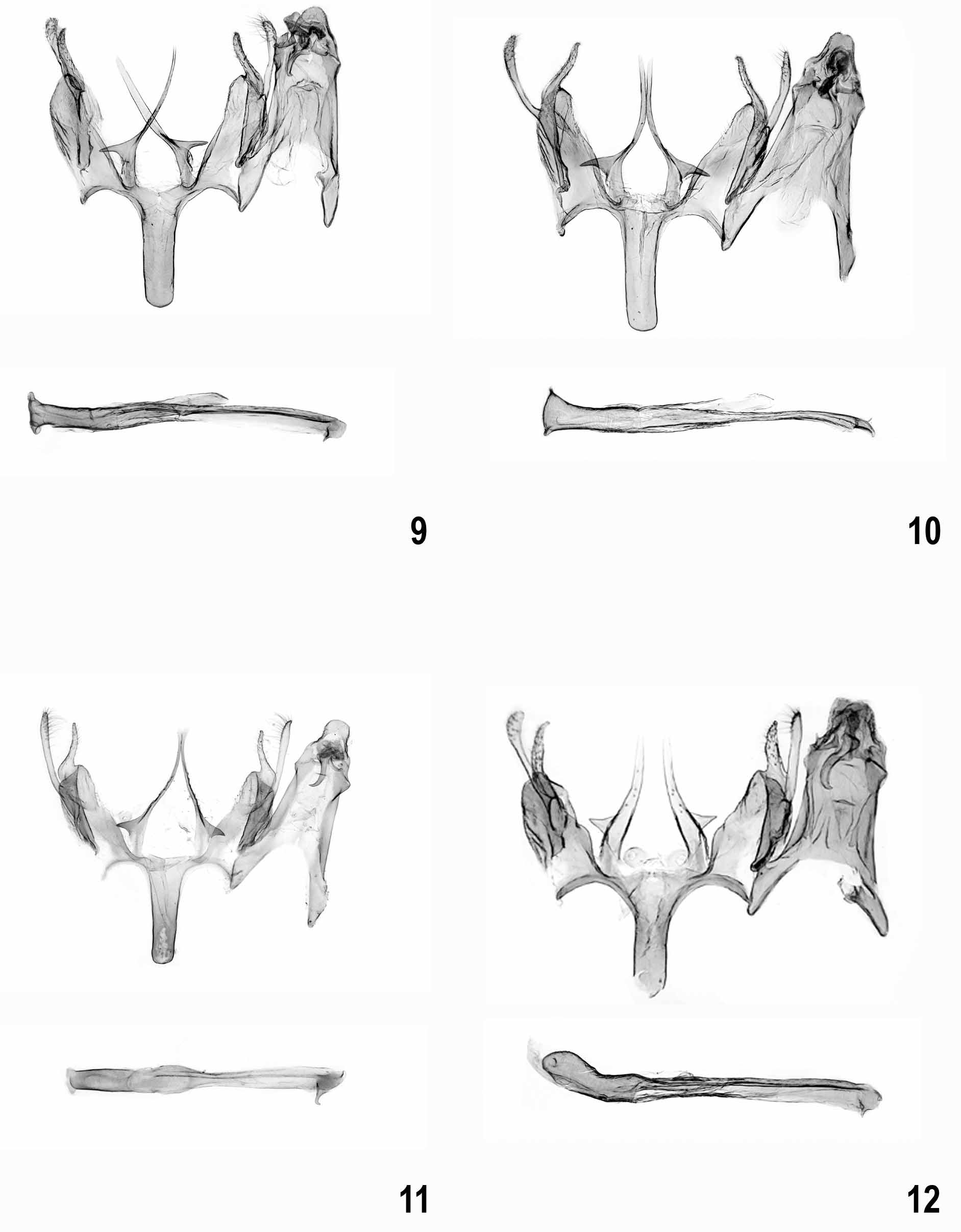
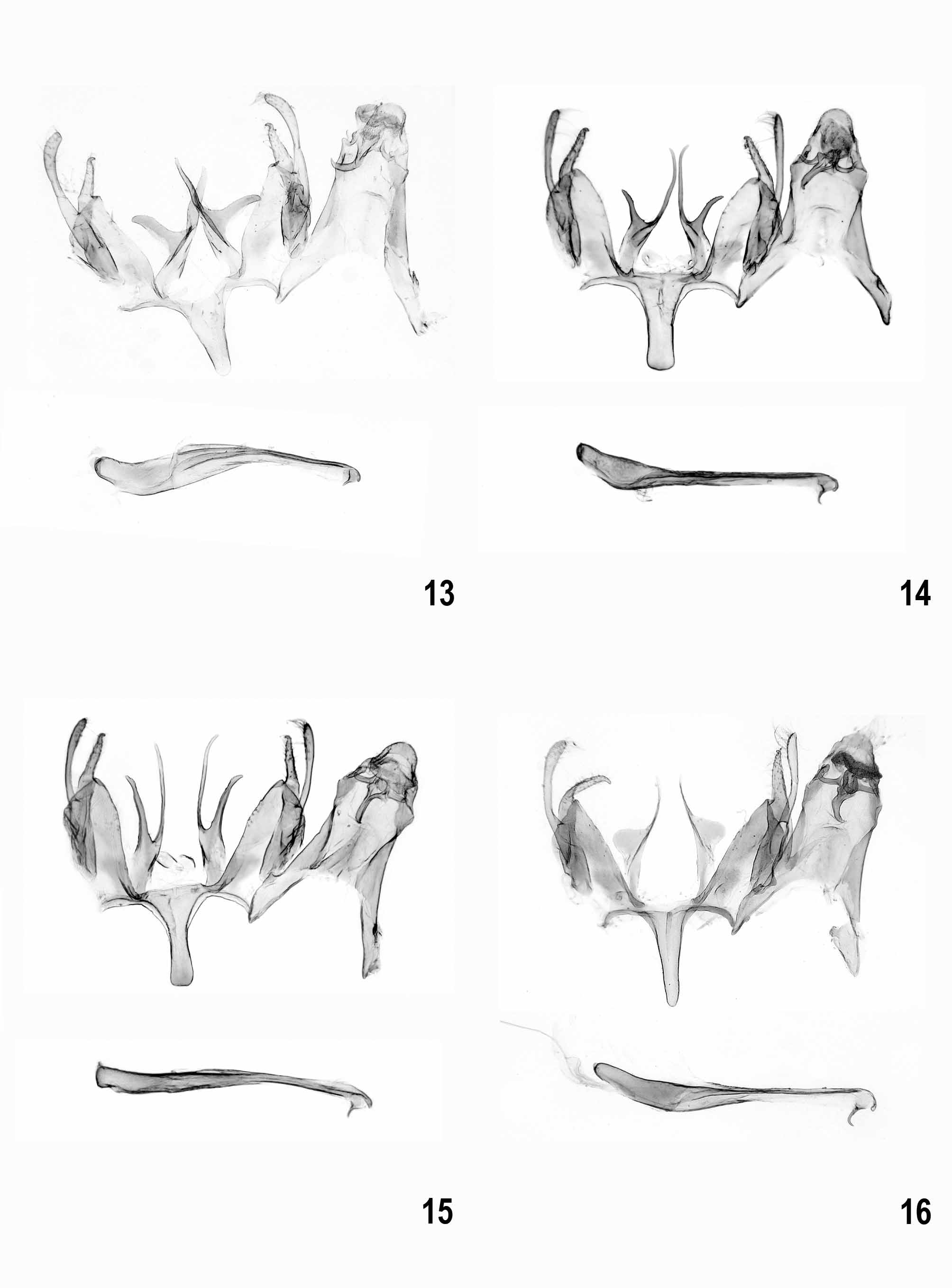
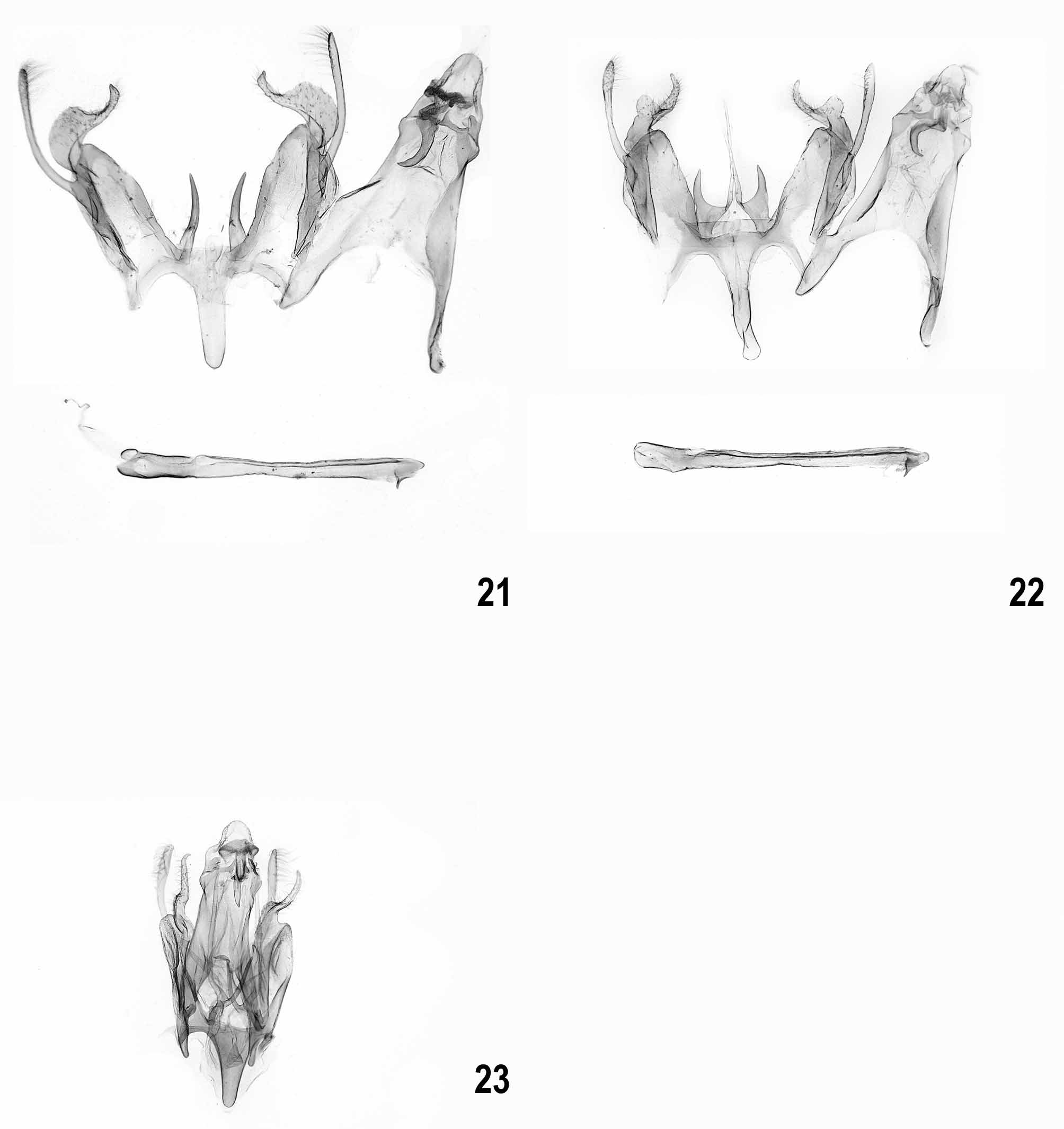
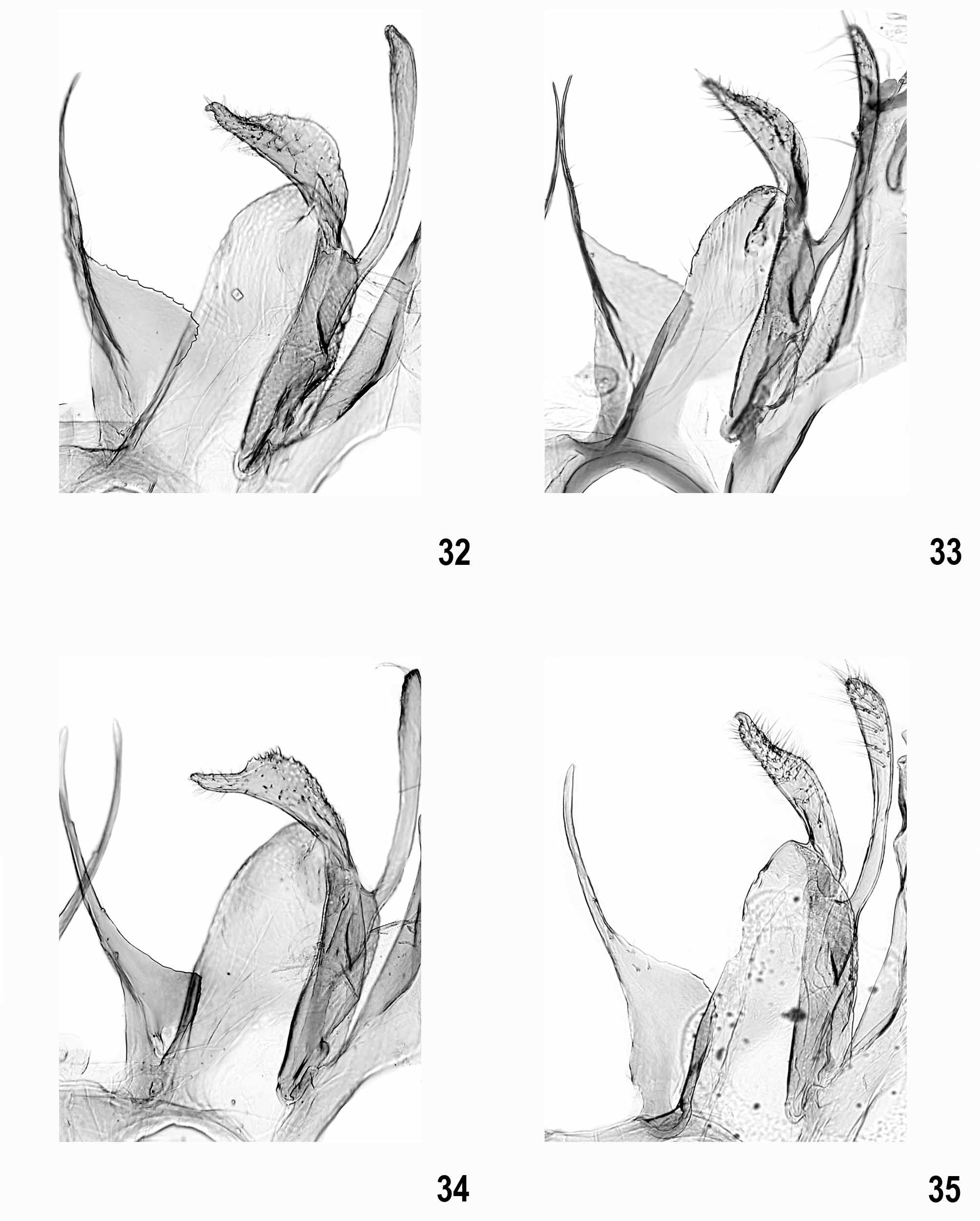
Male genitalia ( Figs 9–35 View FIGURES 9 – 12 View FIGURES 13 – 16 View FIGURES 17 – 20 View FIGURES 21 – 23 View FIGURES 24 – 27 View FIGURES 28 – 31 View FIGURES 32 – 35 ). Subgenital segments: sternite VIII sub-oval, with or without weak emargination medially; tergite VIII tongue-shaped, slender; uncus short and moderately slender, with rounded apex; gnathos hook strong, culcitula distinct; tegumen gradually widened from uncus to pedunculi, anterior margin deeply emarginate; pedunculi long, slender; valva long, slender, straight to weakly curved; sacculus shorter than valva, distally pointed, basal part with or without rounded lobe; vinculum broad, deeply emarginate posteriomedially with pair of long, strongly sclerotized, needle-shaped to forked processes, occasionally with lateral lobe; saccus about length to slightly shorter than processes, broad with almost parallel margins and rounded to cut-off apex; anellus with pair of small, rounded, centrally perforated sclerites; phallus long, slender, straight to weakly curved, coecum weakly separate, apex with sclerotized plate and short arm, ventromedial part occasionally with projection.
Female genitalia (see Huemer and Karsholt 2010). Apophysis posterioris very long; sternum VIII strongly sclerotized, with pair of small and drop-like pockets; sternite VIII medially with irregular folds or ridges; apophysis anterioris about length of segment VIII, about one-third length of apophysis posterioris, basally strongly sclerotized and extended into ostium bursae and to sternal pockets; antrum long and tubular; ductus bursae about length of or shorter than antrum; corpus bursae spherical to oval, usually without, rarely with barely discernible signum.
Bionomics. The biology of most species of Sattleria is insufficiently described and only a few authors such as Burmann (1954) deal with the habits of preimaginal stages. The ovum and oviposition sites are unknown. The larval stages have been recorded from various Caryophyllaceae ( Silene spp., Moehringia sp.) and Saxifragaceae ( Saxifraga spp.). They have been found in July and August, feeding from a loosely spun silken tube that is often extended from the upper parts of the host-plant to the roots or nearby stones ( Pitkin and Sattler 1991). Frass is deposited outside the larval tube. The simultaneous presence of different larval stages indicates a biennial phenology, but phenology may depend on the location and microclimatic conditions. Hibernation probably takes place in the larval stage. Early snowfall shortly after the flight season indicates an alternative hibernation of the pupa or the ovum (Huemer, pers. obs.). The larvae apparently pupate near the ground surface in a dense cocoon, often under stones or amongst the host-plant. Adults have been collected from late June and mid-September. Burmann (1954) observed a mating pair in the early afternoon, whereas Pitkin and Sattler (1991) suspected that increased male activity about midnight was related to their search for females at that time. According to personal observations (P.H.) the males of two species were found in large numbers and extremely active in the early morning at temperatures slightly above zero, indicating mating behaviour. Males of most if not all species are easily attracted to different artificial light sources. Species of Sattleria are restricted to alpine habitats such as scree and rock formations with sparse vegetation as well as alpine grassland, preferably on calcareous, rarely on siliceous soil. The vertical distribution ranges from about 2000 to 3500 m, exceptionally as low as 1500 m.
MAP 1. The distribution of the genus Sattleria in Europe (exclusively based on examined material); altitudinal zones above 1600 m s.l. in blue.
Distribution. Species of Sattleria are restricted to the European mountain systems (Map 1). They have been recorded mainly from the Alps where diversity reaches it highest level with 10 species (Map 2). The genus is also represented in the Pyrénées by three additional species— S. arcuata Pitkin & Sattler, 1991 , S. pyrenaica (Petry, 1904) , and S. angustispina Pitkin & Sattler, 1991 . The Carpathian Mountains are inhabited by a single endemic species, S. dzieduszyckii ( Nowicki, 1864) whereas the Dinaric Mountains and the Apennines share their single species with the Alps ( S. triglavica Povolný, 1987 and S. melaleucella (Constant, 1865) respectively). A record from the Balkan Mountains is dubious and whereabouts of the material are unknown ( Pitkin & Sattler 1991). No records of Sattleria are known from the Caucasus or Mediterranean mountain systems.
Remarks. The external and genitalia morphology for most taxa have been extensively described by Pitkin and Sattler (1991), Povolný (1987, 2001, 2002) and most recently Huemer and Karsholt (2010), and particularly the last mentioned revision may be used as a reference. Descriptions in this publication are restricted to new and revised taxa and because of the absence of females the latter are necessarily omitted from diagnoses.
MAP 2. The distribution of taxa of the Sattleria in the Alps (exclusively based on examined material); altitudinal zones above 1600 m s.l. in blue.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |