
Probreviceps durirostris Loader, Channing, Menegon & Davenport
|
publication ID |
https://doi.org/10.5281/zenodo.172816 |
|
publication LSID |
lsid:zoobank.org:pub:F5906335-FA90-41C8-9679-EED8E34DFF62 |
|
DOI |
https://doi.org/10.5281/zenodo.5696771 |
|
persistent identifier |
https://treatment.plazi.org/id/03CA3B2C-FF96-1336-EE6A-FEE69D39F8FF |
|
treatment provided by |
Plazi |
|
scientific name |
Probreviceps durirostris Loader, Channing, Menegon & Davenport |
| status |
sp. nov. |
Probreviceps durirostris Loader, Channing, Menegon & Davenport View in CoL sp. nov.
Figures 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
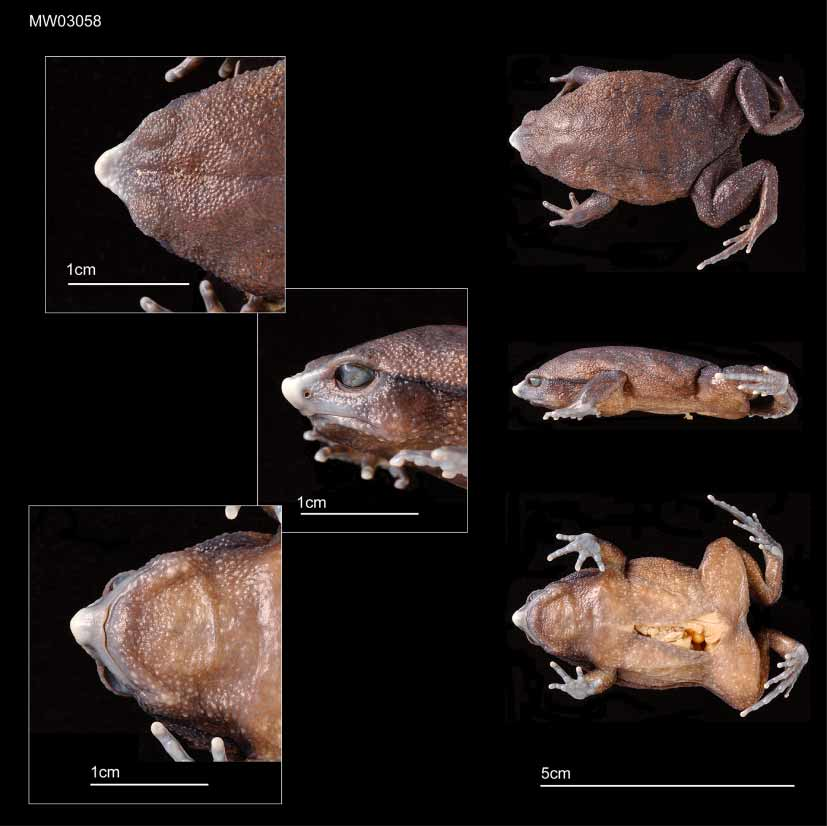
Holotype — A large female, snouturostyle length (SUL) 40.1, BMNH 2005.1564 (field number MW 3058) deposited at the Natural History Museum, London (NHM).
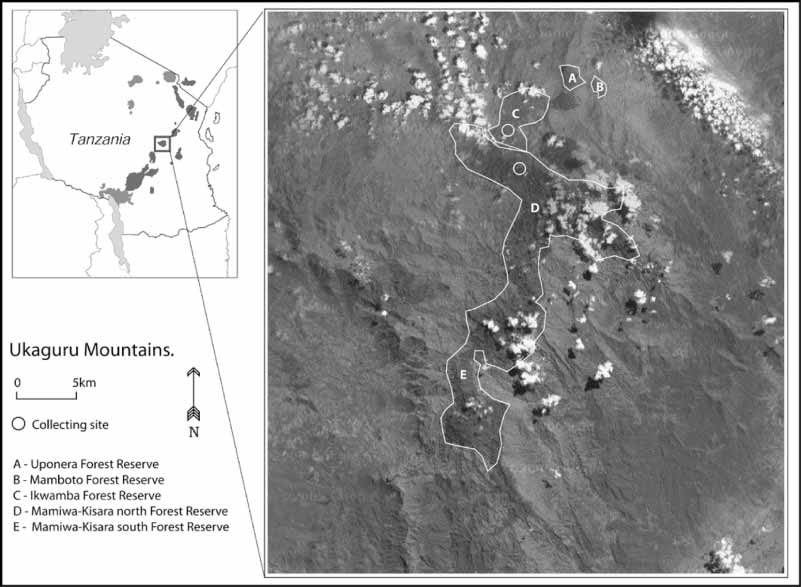
Collected 5 May 2002, by R. Hinde, S. P. Loader and J. Mariaux from Ikwamba Forest Reserve, Morogoro Region, Kilosa District, Ukaguru Mountains, Tanzania, 06º 21’ 31.5” S; 36º 58’ 57.8” E, 1500 m. Tissue has been preserved under the field numbers MW 3059 and 3060.
Paratypes — Eleven specimens, MTSN 554551, collected by M. Menegon and Wilirk Ngalason from MamiwaKisara Forest Reserve, 1850 m, 06° 22’ 48’’S, 36° 56’ 02’’ E, deposited at Museo Tridentino di Scienze Naturali, Trento, Italy ( MTSN 5547 is now accessioned in the NHM, as BMNH 2005.1573 and is cleared and stained. Tissue samples MW 4089–90 were taken from MTSN 5546–47 respectively); and four specimens MHNG 2670.32, MHNG 2670.33, MHNG 2670.31 and UDSM 2326 were collected by William T. Stanley from “Camp A”, which is in slightly degraded primary forest, and has the locality: Tanzania, Morogoro Region, Kilosa District, Ukaguru Mts, MamiwaKisara Forest Reserve, 1 km E, 0.75 km S Mt. Munyera, 1900 m, 06° 22’ 45” S, 36° 56’ 10” E.
Diagnosis
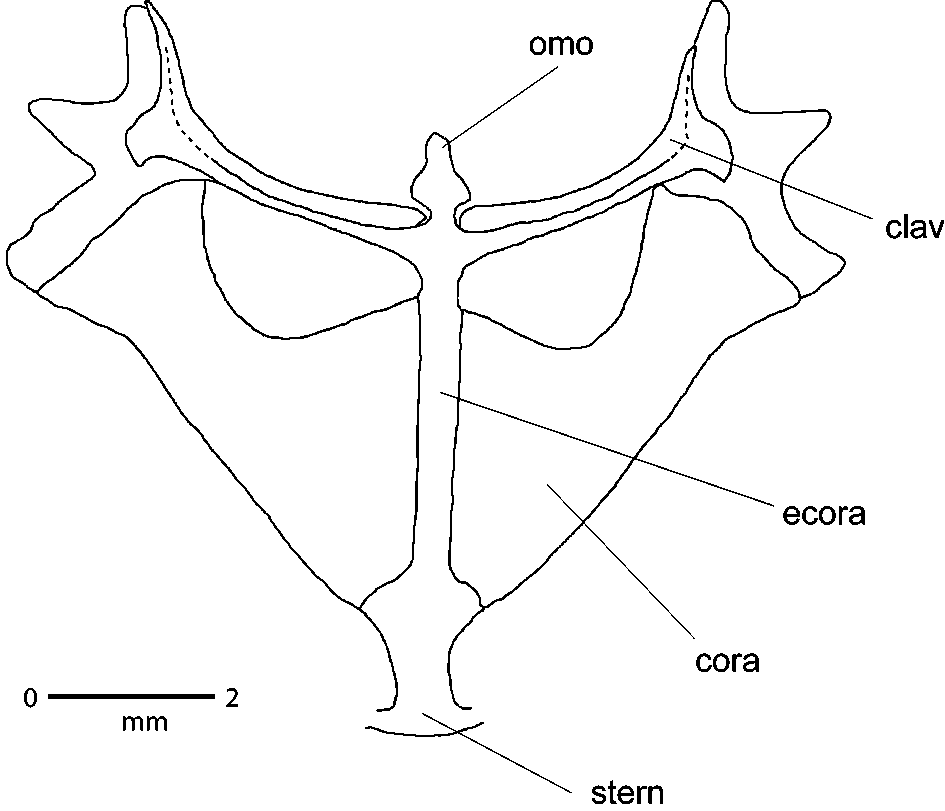
The species is a brevicipitid on account of the following combination of characters, mainly associated with the retention of the shoulder girdle ( Parker, 1934, see Fig. 3 View FIGURE 3 ): (1) Omosternum cartilaginous (2) Fully developed clavicles and procoracoid cartilages (3) Dilated coracoids (4) Cartilaginous sternum (5) Expanded vomer. The species is assigned to the genus Probreviceps based on the following characters, which separate it from other brevicipitines: (1) Fusion of the urostyle and the sacral vertebra (unfused in Callulina and Spelaeophryne ). (2) Palate smooth with two serrated denticulate ridges sensu Parker, (1934); Largen and Drewes, (1989) ( Fig. 4 View FIGURE 4 ) (absent in Breviceps , single row in Callulina , Spelaeophryne , Balebreviceps ). (3) Omosternum moderately large (reduced/vestigal in Breviceps and Spelaeophryne ) (4) Moderately dilated coracoids (as in Spelaeophryne and Callulina but strongly in Balebreviceps , Breviceps ) (5) Terminal phalange simple (expanded in Callulina ).
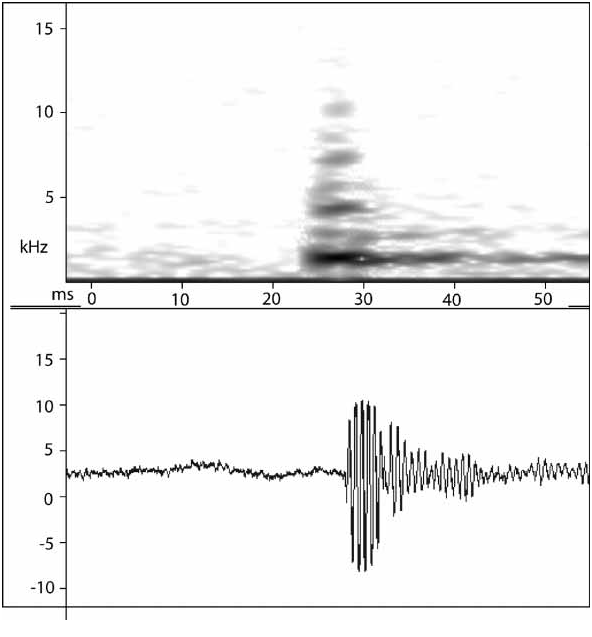
P. durirostris is a medium sized Probreviceps which can be easily distinguished from all other species: (1) Snout morphology: P. durirostris has a markedly pointed snout, which is hardened white with ridges along the canthus to the apex of the snout (see Fig. 1 View FIGURE 1 ). Probreviceps m. rungwensis has a pointed snout, but is not hardened, white in colour, nor does it have a ridged canthus. All other species of Probreviceps have relatively rounded snouts. (2) Tympanum size: P. durirostris is sexually dimorphic with respect to tympanum size, which varies from 13 to 16% of SUL in males, and 4 to 7% in females. This is similar to P. m. rungwensis, which varies from 9 to 14% in males, and 4 to 7% in females. Both P. m. loveridgei and P. m. macrodactylus males do not possess such large tympani, varying from 4 to 7% in males, and 3 to 5% in females. Four P. rhodesianus specimens measured vary in typmanum size 3–5% of SUL. P. uluguruensis does not have a visible tympanum. (3) Finger ridging: Parker reported a fleshy ridge along the inner side of the second and third finger in P. uluguruensis . Finger ridging is present in P. durirostris , though in some specimen only faintly and not as strongly as P. uluguruensis . Similarly, in P. m. rungwensis and P. rhodesianus finger ridging is present, though very slight in the latter. The ridge is almost entirely absent in P. m. macrodactylus and P. m. loveridgei . (4) Position of vent: The vent opens posteroventrally in P. durirostris (as in P. m. macrodactylus , P. m. loveridgei , and P. m. rungwensis) (opens ventrally in P. rhodesianus , and P. uluguruensis ). (5) Advertisement call: the new species can be distinguished from all other species recorded. The call of P. durirostris is a very slow series of clicks, slightly faster than one every two seconds ( Fig.5 View FIGURE 5 ). This contrasts with the calls of three taxa of Probreviceps in Tanzania that are rapidly pulsed chirps ( Mkonyi et al 2004), varying from a mean of 20/sec ( P. uluguruensis ); 38/sec ( P. m. macrodactylus ) to 90/sec ( P. m. loveridgei ). The calls of P. m. rungwensis from Mount Rungwe and the Udzungwa Mountains in Tanzania, and P. rhodesianus from Zimbabwe, are unknown.
Description of holotype
The SUL is 34.3mm. The head is flat on top with a protruding snout, 5.0mm, the canthal ridges, tip of snout and the upper lip are white and hardened ( Fig 1 View FIGURE 1 ). The canthal ridges, which form an apex at the anterior of the snout are continuous with the ridged skin of the eyelids. The apex of the snout is ridged and slightly raised and pointed in profile. Loreal region is almost vertical, slightly concave. Snout tip to mouth 3.0mm. Maximum head width 14.0mm. Nostrils are round, situated midway between lip and top of snout, with the openings visible laterally. Tongue longer than wider, posterior half and lateral edges not adhered to the lower jaw. Choanae are small and round. Internostril distance 3.3. Snout tip to nostril 2.2mm, and nostril to eye 2.5mm. The eyelid ( 4.7mm) extends back as a strong supratympanic ridge to above the arm as a fold in the dorsal skin. The eyes are visible from below, eye 3.6mm. Interorbital distance measured across the top of the head 4.3mm. The anterior corners of the eyes are 6.2mm apart. The tympanum is round, 2.8mm, placed 2.5mm from eye, and aligned vertically when viewed from the front. The tubercles of the hand are pale, large with faint skin ridges (fig 1). First finger is the shortest, with second and fourth toe slightly larger, the third being the longest. Length of third finger to include basal tubercle 7.25mm. The legs are relatively short, with a tibia length 17.2mm. First toe is the shortest, with second and fifth toes slightly longer, roughly similar in length, third toe is nearly twice as long as second and fifth, with the fourth the longest. The length of the fourth toe ( Fig 1 View FIGURE 1 ), including the outer metatarsal tubercle 18.6mm. The subarticular tubercles of the toes are rounded, pale, with faint skin ridges. Inner metatarsal tubercle well developed, rounded, length 2.9mm, while outer is large but flattened. Skin rough with small distinct glands that tend to merge on the lower back and upper legs. They are small anteriorly, becoming bigger posteriorly. The only parts not covered with glands are the plantar surfaces of the hands and feet. The vent opens posteroventrally. The following are the body proportions: Maximum head width/SUL 0.35, snout/eye 1.39, eye/ SUL 0.09, eyetympanum distance/horizontal tympanum 0.89, tympanum/SUL 0.07, anterior corners of eyes/head width 0.44, snout tip to mouth/internostril distance 0.91, internostril/nostrileye 1.32, third finger/head width 0.52, tibia/head width 1.23, tibia/SUL 0.43, fourth toe/SUL 0.40, inner metatarsal tubercle/fourth toe 0.16.
Colour
In life: The back and sides have dark warts on a tan brown background. Five faint transverse pairs of darkgrey narrow bands are evenly spaced on the posterior dorsum, angled diagonally and posteriorly from the midline. Small whitetipped warts are irregularly present on dorsum and upper limb surfaces. The snout tip is pale bluegrey, with the side of the head dark grey to black from side of the snout to the arm insertion, running below the upper eyelid and the supratympanic fold. Pupil horizontal with a gold iris. The tympanum is dark grey to black. The upper surfaces of the limbs are dark grey ( Fig 2 View FIGURE 2 ).
In alcohol: The background dorsal colour is brown, with darker warts. Four pairs of darker transverse lines are just visible, from midway down the back to the legs. Glands are present on the belly. Back of ankle, and underside of forearm are white.
Paratype variation
The paratypes are similar to the holotype in proportions and colour. Males vary in SUL from 25.8 to 26.8, with the females larger at 34.3 to 43.1. The males have large tympani, 13 to 16% of SUL, but 4 to 7% in females. Ranges of body proportions for the 11 paratypes include: Maximum head width/SUL 0.34–0.49, snout/eye 0.88–1.35, eye/SUL 0.08–0.13, eyetympanum distance/horizontal tympanum 0.20–0.31 (males), 1.11–1.86 (females), tympanum/SUL 0.13–0.16 (males), 0.04–0.07 (females), anterior corners of eyes/head width 0.39–0.47, snout tip to mouth/internostril distance 0.65–0.94, internostril/ nostril–eye 1.05–1.45, third finger/head width 0.36–0.73, tibia/head width 0.88–1.30, Probreviceps durirostristibia/SUL 0.39–0.46, fourth toe/SUL 0.37–0.46, inner metatarsal tubercle/fourth toe 0.10–0.17. Finger ridges vary from prominent to faint.
Advertisement call
Males call actively during the day, with some residual calling at night. Calling takes place from within leaf litter or while the animals are concealed under logs or other forest debris. Choruses form rapidly after an individual starts calling. Males were recorded in the field at the MamiwaKisara Forest Reserve. The call is a slow series of clicks ( Fig 5 View FIGURE 5 ), produced at a rate of approximately 32/minute. Each click consists of a single highenergy pulse with a duration of 6 ms. No real harmonics are present, although significant energy is present at 1.2, 1.6, 2.6, 4.3 and 5.4 kHz.
Phylogenetic analyses
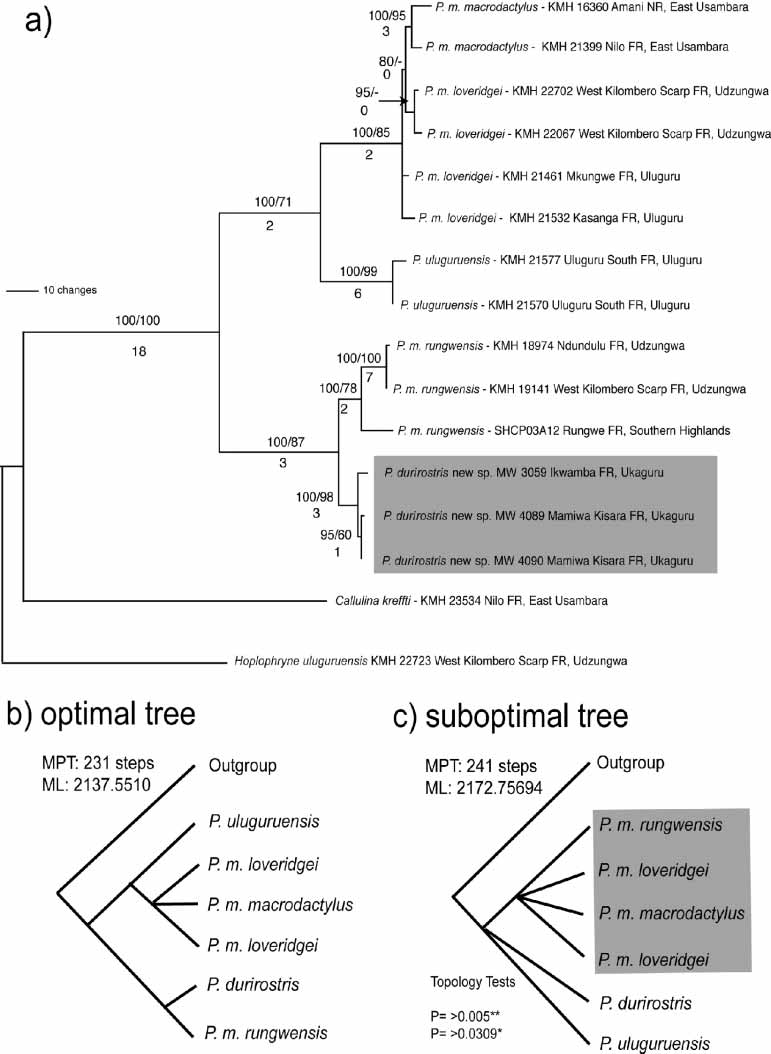
Alignment of sixteen DNA sequences (concatenated 12S and 16S sequences) resulted in a matrix of 717 unambiguously aligned characters, of which 557 were constant and 160 variable; of these, 81 were informative under parsimony. Fourteen Probreviceps were included in the analysis, one brevicipitine Callulina , and the microhylid Hoplophryne uluguruensis was used as an outgroup, based on its position outside of a brevicipitine clade in previous analyses (Loader et al. 2004). Maximum likelihood (heuristic search using 10 randomaddition sequence replicates and TBR swapping) and Bayesian analyses were carried out under a GTR I + G model as calculated by Modeltest 3.04 ( Posada and Crandall, 1998). Base frequencies were estimated as 0.3143, 0.2341, 0.2241, and 0.2275 for A, C, G and T respectively, substitution rates = 8.6657 26.7726 19.7332 3.2566 77.3197 with the proportion of invariant sites set at 0.4201 and a gamma distribution shape parameter 0.6367. The phylogeny recovered is shown in Fig. 6a View FIGURE 6. a . An exhaustive search option using parsimony yielded eight best trees (231 steps), identical to the phylogeny recovered in ML and Bayesian analyses, differing only in the resolution of P. m. macrodactylus , and P. m. loveridgei from Uluguru, Usambara and Udzungwa which collapse into a polytomy in the consensus tree. Support for clades was measured with P. durirostrisbootstrap proportions for 1000 pseudoreplicates ( Felsenstein, 1985), Bayesian posterior probability, and decay indices ( Bremer, 1988). In summary, the tree demonstrates that Probreviceps durirostris is monophyletic, and is the sister species to Probreviceps m. rungwensis. Probreviceps m. rungwensis is also shown to have two divergent lineages, which each correspond to distinct areas the Udzungwa and Southern Highlands. For further details of the analysis of species and intrageneric relationships in brevicipitines, see Loader et al. (2004).
One alternative tree arrangement was investigated to evaluate a previous taxonomic hypothesis based on morphology ( Parker, 1934), i.e. the monophyly of P. macrodactylus . Using parsimony, a Templeton test ( Templeton, 1983) compared the optimal tree ( Fig. 6 View FIGURE 6. a b) with constrained suboptimal topology ( Fig. 6 View FIGURE 6. a c). Likelihood tests (ShimadairoHasegawa Test) were similarly applied to compare optimal and suboptimal trees ( Fig. 6 View FIGURE 6. a c). The best suboptimal trees including a constrained monophyletic P. macrodactylus subspecies complex (see Fig. 6 View FIGURE 6. a c), are a significantly worse fit to the data than the optimal arrangements. This result indicates, as shown by Loader et al. (2004), that P. macrodactylus requires reevaluation, the subject of a paper in preparation.
Distribution
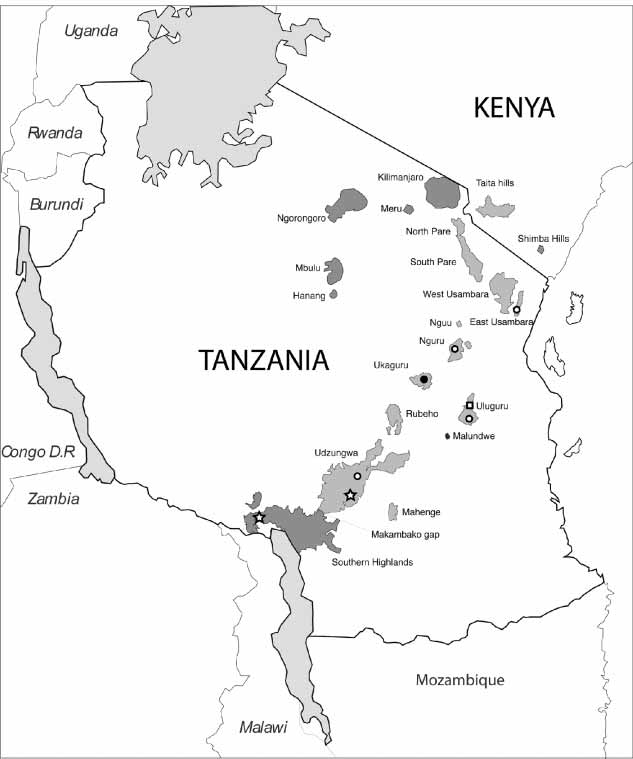
This new species is presently only known from two forest reserves in the Ukaguru Mountains; MamiwaKisara Forest Reserve and the Ikwamba Forest Reserve ( Fig 7 View FIGURE 7 ).
Etymology
The name durirostris is derived from the latin “dura” meaning hardened and “rostris” meaning nose. The name is used in reference to the distinctive hardened nose of the new species.
Remarks
The new species P. durirostris resembles P. m. rungwensis in general body shape, including the enlarged snout. Their morphological similarity is mirrored in their close phylogenetic relationship as recovered in the optimal trees. It is unclear whether the pointed snout has any specific function, for example, the snout might be used to facilitate excavation of burrows. Probreviceps are believed to primarily burrow using their shovel like hind limbs, being backward burrowers ( Van Dijk, 2001). Investigation of the behavioural ecology of each Probreviceps species (with and without pointed noses), and their associated habitats will need to be carried out to infer any functional associations.
The distribution of Tanzanian Probreviceps species and subspecies is shown in Fig. 8 View FIGURE 8 .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |