
Parahelichus granulosus ( Delève, 1974 ), Deleve, 1974
|
publication ID |
https://doi.org/10.11646/zootaxa.3852.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:033A771C-839D-41C5-91C3-7D02A661F4C9 |
|
DOI |
https://doi.org/10.5281/zenodo.5630235 |
|
persistent identifier |
https://treatment.plazi.org/id/03C91474-FFFC-8E07-FF09-7907FAF89BFC |
|
treatment provided by |
Plazi |
|
scientific name |
Parahelichus granulosus ( Delève, 1974 ) |
| status |
|
Parahelichus granulosus ( Delève, 1974)
Helichus granulosus Delève 1974: 171
Taxonomy. Delève (1974) described this species in the genus Helichus View in CoL , providing the following comment on its placement into the subgenus Parahelichus View in CoL : “Par la forme du pronotum cette espèce pourrait rentrer dans le sousgenre Parahelichu s Bollow (1940) mais le corps est dépourvu de la pubescence serrée masquant la coloration du tegument, caractère ayant servi à fonder le sous-genre.” The main difference between the subgenera Praehelichus View in CoL and Parahelichus View in CoL is in the form of lateral pronotal carina that is regularly bent in Praehelichus View in CoL , while unevenly bent or lobate in Parahelichus View in CoL . Beside this key feature Bollow (1940) mentioned the 10-segmented antennae for Parahelichus View in CoL and 11-segmented for Praehelichus View in CoL . Although the classification of these two supraspecific taxa has not yet been sufficiently resolved, we consider P. granulosus and P. pseudogranulosus to form a distinct species group within the genus based on the shape of pronotal carina and tightly arranged small granules on the surface.
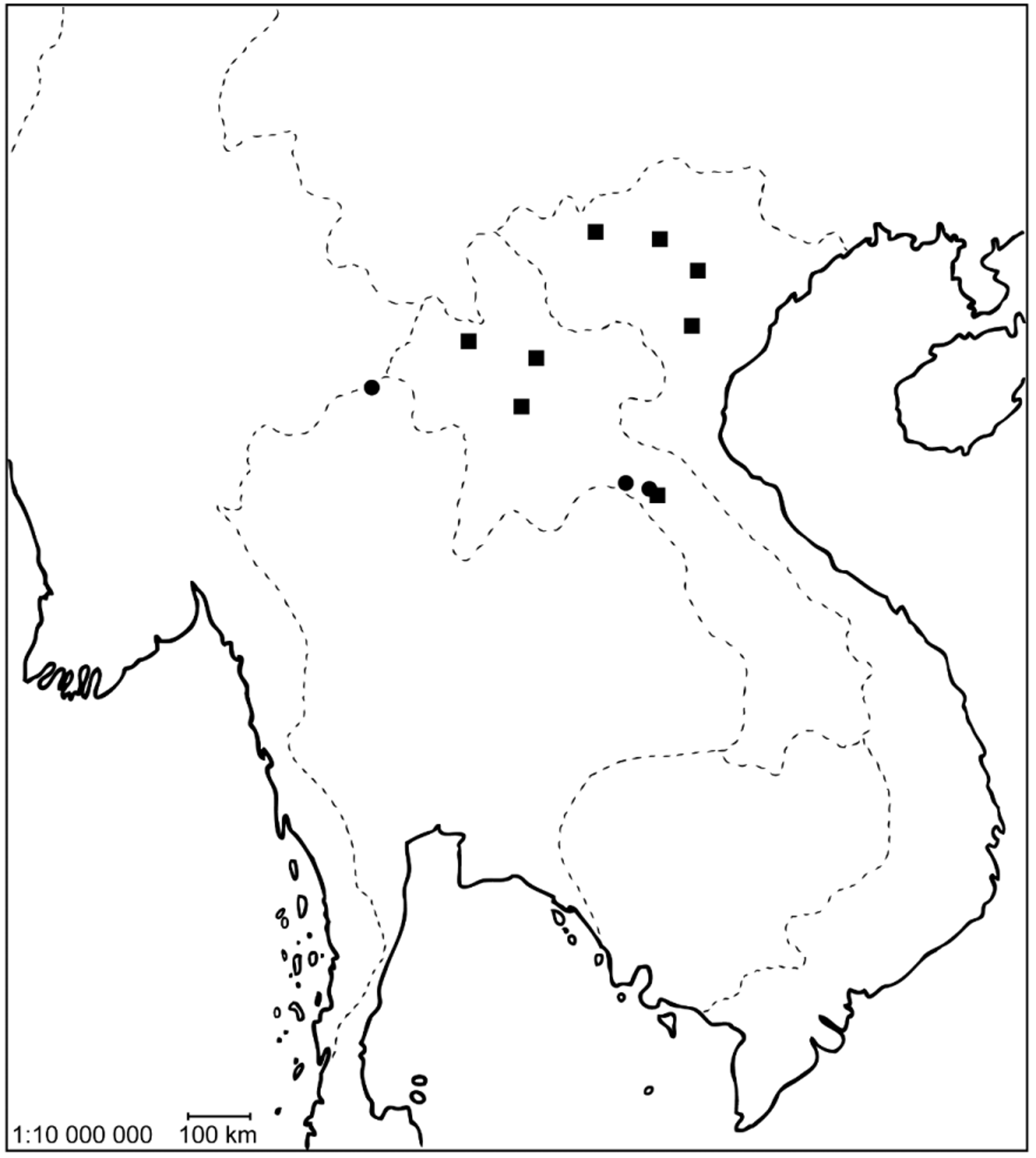
Type locality. Hanoi, Vietnam, single holotype specimen was collected by light in hotel Kim lien ( Delève 1974).
Material examined. Holotype ♂ ( MTMB): “Hanoi, Hotel Kim lien, 28. 31-V- 1966 Gy. Topál, n-588, collected by lamp.” Additional material. 442 ex. ( CKB, MTMB, NMW, NHGH): 16 ♂♂, 12 ♀♀: “ Vietnam, Yen Bai, 15.V. 1990, J. Horák leg.”; 10 ♂♂, 10 ♀♀: “ Vietnam, Yen Bai, 6.–25. 5. 1990, O. Šauša lgt.”; 1 ♂: “ Vietnam, Vinh phu prov., Tam dao, 27.5.– 2.6. 1986, Jan Horák lgt.”; 3 ♀♀: “ Vietnam, Ha son bin prov., Hoa binh, 4.– 7.6. 1986, Jan Horák lgt.”; 91 ♂♂, 92 ♀♀: “N - Vietnam, Sa Pa (Lao Cai), 25.5.– 10.6. 1991, 22° 20`N, 103° 50`E, E. Jendek leg.”; 25 ♂♂, 17 ♀♀: “ Laos north, Louang Namtha, 20km NW, N 21° 0 9.2, E 101° 18.7, alt. 900 ± 100 m, 5.–11. V. 1997, E. Jendek & O. Šauša leg.”; 70 ♂♂, 86 ♀♀: “ Laos - N, Louangphrabang pr. 19° 53`N, 102° 09`E, Khan riv., 300 m, 21.IV. 1999, Vít Kubáň leg.”; 1 ♂: “ Laos, Louangphrabang pr. 20° 33-4`N, 102° 14`E, Ban Song Cha ( 5 km W), 1200 m, 1.–16. V. 1999, Vít Kubáň leg.”; 3 ♂♂, 5 ♀♀: “ Laos, centr., Khammouan prov., 18° 07`N, 104° 29`E, alt. 250 m, Ban Khoun Ngeun env., 10.–13. VI. 2001, E. Jendek & O. Šauša leg.”.
Differential diagnosis. P. granulosus and P. pseudogranulosus sp. nov. are large species distinguished from all described Asian Praehelichus , Parahelichus and Helichus species by the very tightly arranged small granules on the dorsal and ventral surface (causing its reticulate appearance at smaller magnifications). Further, both species differ from other relatives by the lack of a conspicuous area on the dorsal surface covered by plastron setae. Thus, plastron setae are confined only to a small area on ventrite 1 posteriad of metacoxae in P. granulosus and P. pseudogranulosus , while the remaining congeners bear more or less densely arranged plastron setae on the dorsal surface.
Redescription. Habitus of male as shown in Fig. 1. Body elongated, heavily sclerotized and compact, moderately convex dorsally. Length (BL) in ♂♂: 3.58–6.39 mm (5.36 ± 0.48 mm), in ♀♀: 3.97–6.95 mm (5.84 ± 0.46); maximum width (EW) in ♂♂: 1.50–2.63 mm (2.21 ± 0.19 mm), in ♀♀: 1.61–2.95 mm (2.46 ± 0.19 mm). Colour nearly black, antennal clubs, mouthparts and tarsi reddish-brown; immature specimens brownish.
Surface of pronotum, elytra and venter opaque, covered with tightly arranged round small granules and larger granules. Small granules about one third of a diameter of a facet. Larger granules shiny, nearly rounded, subequal with a diameter of a facet, associated with moderately long yellowish setae and more or less regularly intermixed within small granules. Elytra lacking distinct striae or intervals.
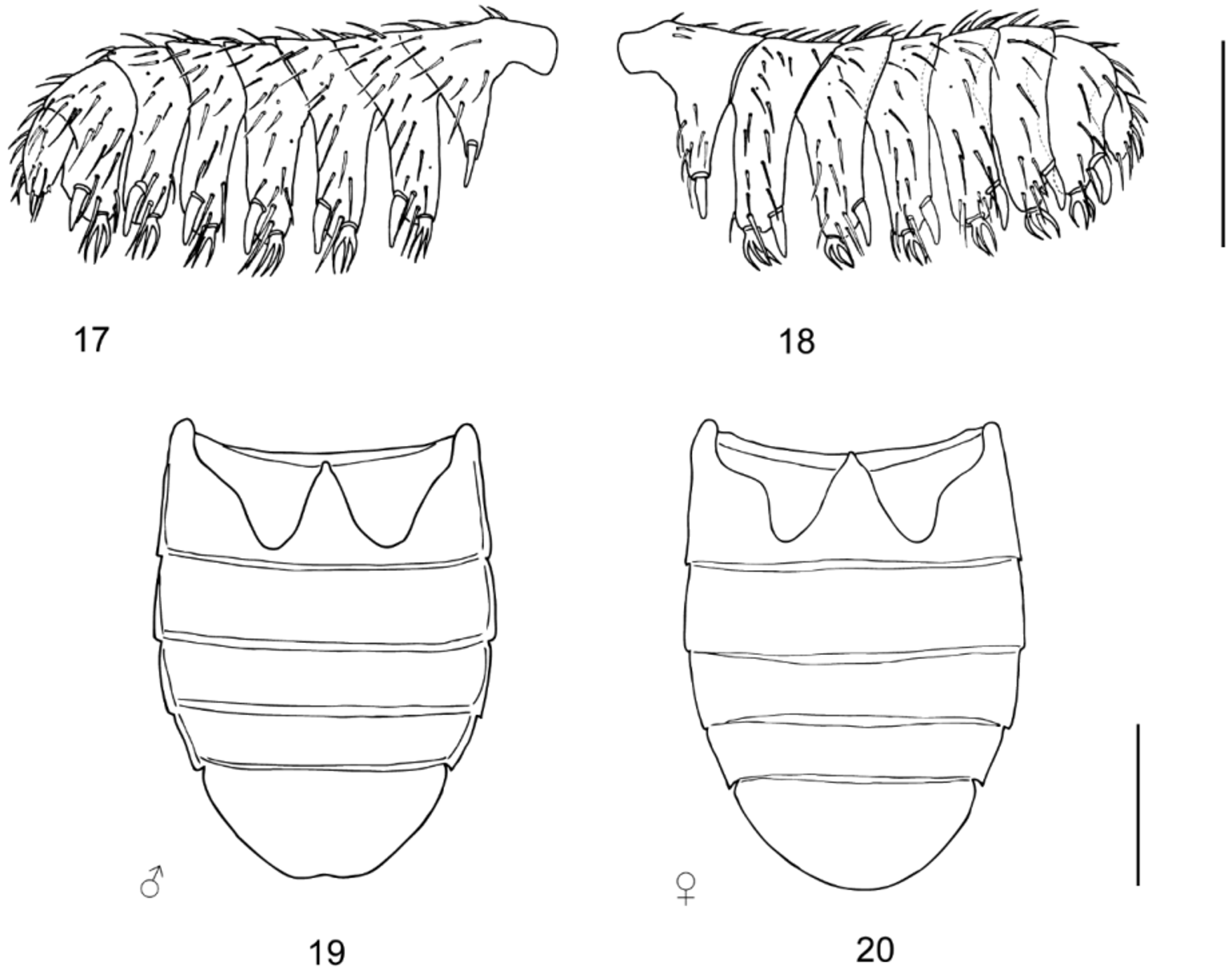
Head wider than long; surface shiny with sparse flat granules; granules moderately smaller than a facet diameter, with yellowish recumbent setae, more densely arranged on anterolateral portion than mesally. Frons with narrow, small shiny longitudinal area; eyes large and protuberant, strongly facetted; interfacetal setae fine, short, scattered and present on central portion of eye. ID in ♂♂: 0.34–0.68 mm (0.58 ± 0.06 mm), in ♀♀: 0.42–0.82 mm (0.66 ± 0.06 mm); HW in ♂♂: 0.76–1.26 mm (1.10 ± 0.09 mm), in ♀♀: 0.87–1.42 mm (1.21 ± 0.09 mm). Antennal insertion exposed from above, antennae widely separated; subantennal groove shallow, reaching about middle of eye; antenna pectinate, short, club with eight antennomeres ( Figs 17, 18 View FIGURES 17 – 20 ).
Thorax. Pronotum wider than long; disc convex dorsally, without gibbosities, carinae or furrows; granules more distinct than those on head, separated usually by distance of one granule diameter or less. Sides arcuate; lateral pronotal carina distinct, moderately raised, not crenulate; posterior angles almost rectangular, moderately acute; anterior angles acute, protruding; posterior margin trisinuate, smooth; prescutellar foveae absent. MPL in ♂♂: 0.89–1.55 mm (1.22 ± 0.10 mm), in ♀♀: 0.92–1.55 mm (1.34 ± 0.10 mm); APW in ♂♂: 0.87–1.42 mm (1.22 ± 0.10 mm), in ♀♀: 0.89–1.55 mm (1.33 ± 0.10 mm); PPW in ♂♂: 1.16–2.00 mm (1.69 ± 0.15 mm), in ♀♀: 1.29–2.24 mm (1.84 ± 0.16 mm). Hypomeron broadest near middle, distinctly narrowed anteriorly and slightly posteriorly; separated from prosternum by a gap along anterior fourth; postcoxal process very short, triangular, not reach prosternal process. Prosternum in front of coxae approximately as long as prosternal process, anterolateral portion rounded. Setae in middle of prosternum and on prosternal process longer than those on dorsal surface; longest setae in disc of metaventrite. Prosternal process wide, strongly overlapping mesoventrite, fitting into deep cavity of mesoventrite, apex long and subacute, sides raised. Mesoventrite very short, with conspicuous ventral projection on each side of mesoventral cavity; exposed portion of mesanepisternum large, about as long as width of hypomeron; mesepimeron fourth as long as metanepisternum, both divided by fine line and both reaching mesocoxal cavity. Mesocoxal cavities subcircular, transversally oriented; mesocoxae subglobular, with short projection near attachment of trochanter, less widely separated than procoxae; exposed portion of mesotrochantin small. Scutellum (scutellar shield) large, broadly rounded, in same level with elytra, surface granulated. Mesoventral and metaventral processes solidly fused. Metaventrite distinctly longer than prosternum in front of coxae, without grooves or depressions for reception of femur and tibia; discrimen long and fine; transverse suture fine but distinct; anterior edge of metaventrite without transverse carina between mesocoxal cavities; metanepisternum wide and long. Metacoxal cavities large, transverse, oblique and not reaching elytra. Metacoxae separated by suture from metaventrite, posteriorly deeply excavate for reception of femora; moderately widely separated mesally, posterior projection distinct. Elytra convex dorsally, lateral margin not explanate, gradually extended from shoulders up to 0.75 of their length, elytral apices subacute; granulation on elytra similar to that on pronotum but large granules less dense and nearly regularly spaced on disc, denser laterally; epipleura widest basally, inflected at level of metacoxa and near apex, partly concealed by lateral margin of ventrites. EW in ♂♂: 1.50–2.63 mm (2.21 ± 0.19 mm), in ♀♀: 1.61–2.95 mm (2.46 ± 0.19 mm); EL in ♂♂: 2.68–4.89 mm (4.14 ± 0.39 mm), in ♀♀: 3.05–5.47 mm (4.50 ± 0.38 mm). Legs slightly longer than elytra; trochanters large, femoral attachment oblique. Femora excavate for reception of tibiae, strongly widened, widest near middle, with scattered fine punctures; mesofemora without rows of conspicuous strong setae on posterior face. Tibiae slightly longer than femora; mesotibia as long as protibia, apically moderately widened; metatibia longest, apically strongly widened; each tibia weakly curved, narrow, with various specialized setae along anterior (protibia) or posterior face (meso- and metatibia) and with numerous short setae on remaining parts. Tarsi approximately as long as protibia and mesotibia, respectively, and slightly shorter than metatibia, with few short strong setae ventrally and longer setae mainly dorsally and laterally; terminal tarsomere on metatibia slightly shorter than combined lengths of tarsomeres 1–4, those on protibia and mesotibia approximately as long as combined lengths of tarsomeres 1–4. Claws robust and simple; empodium squamiform.
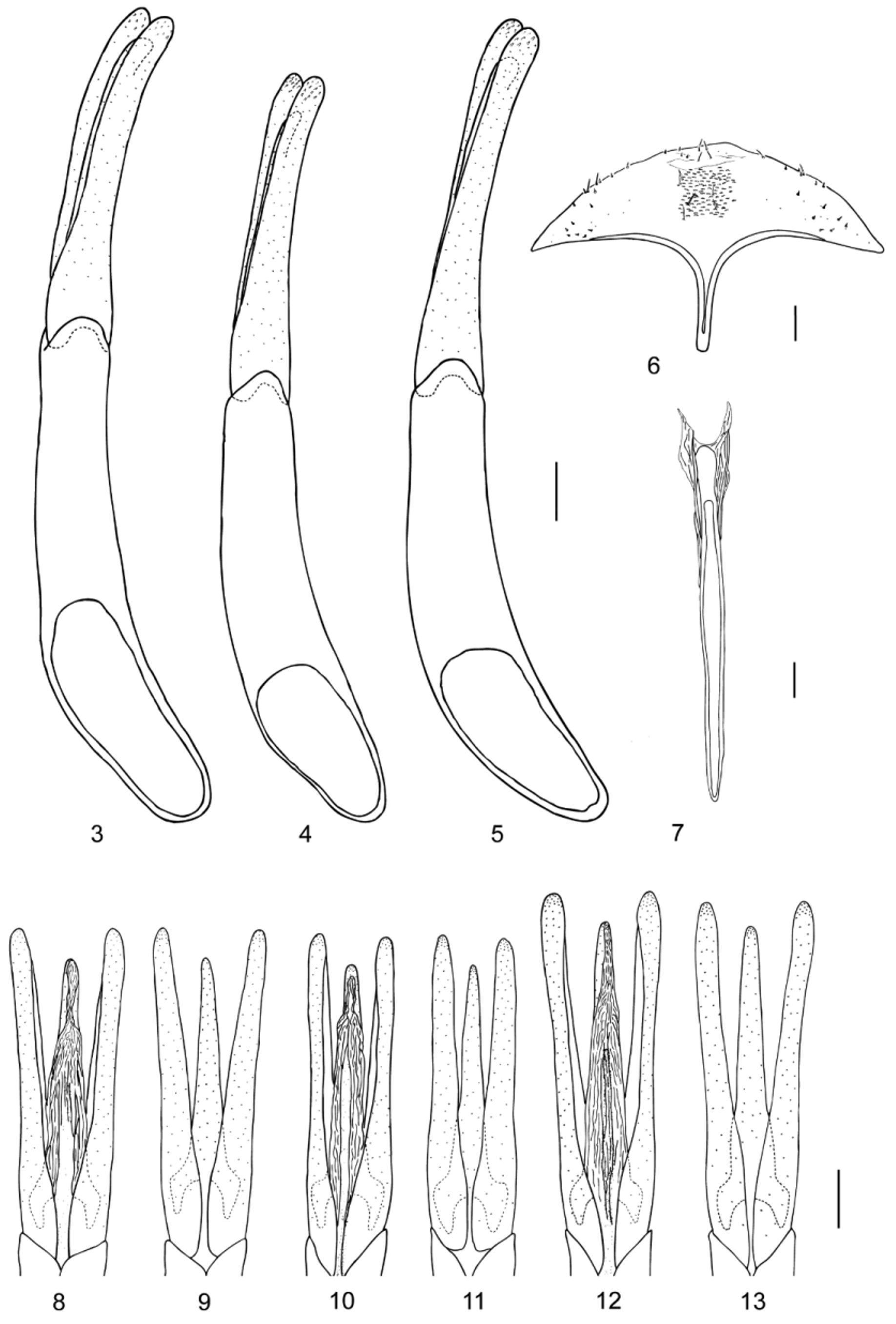
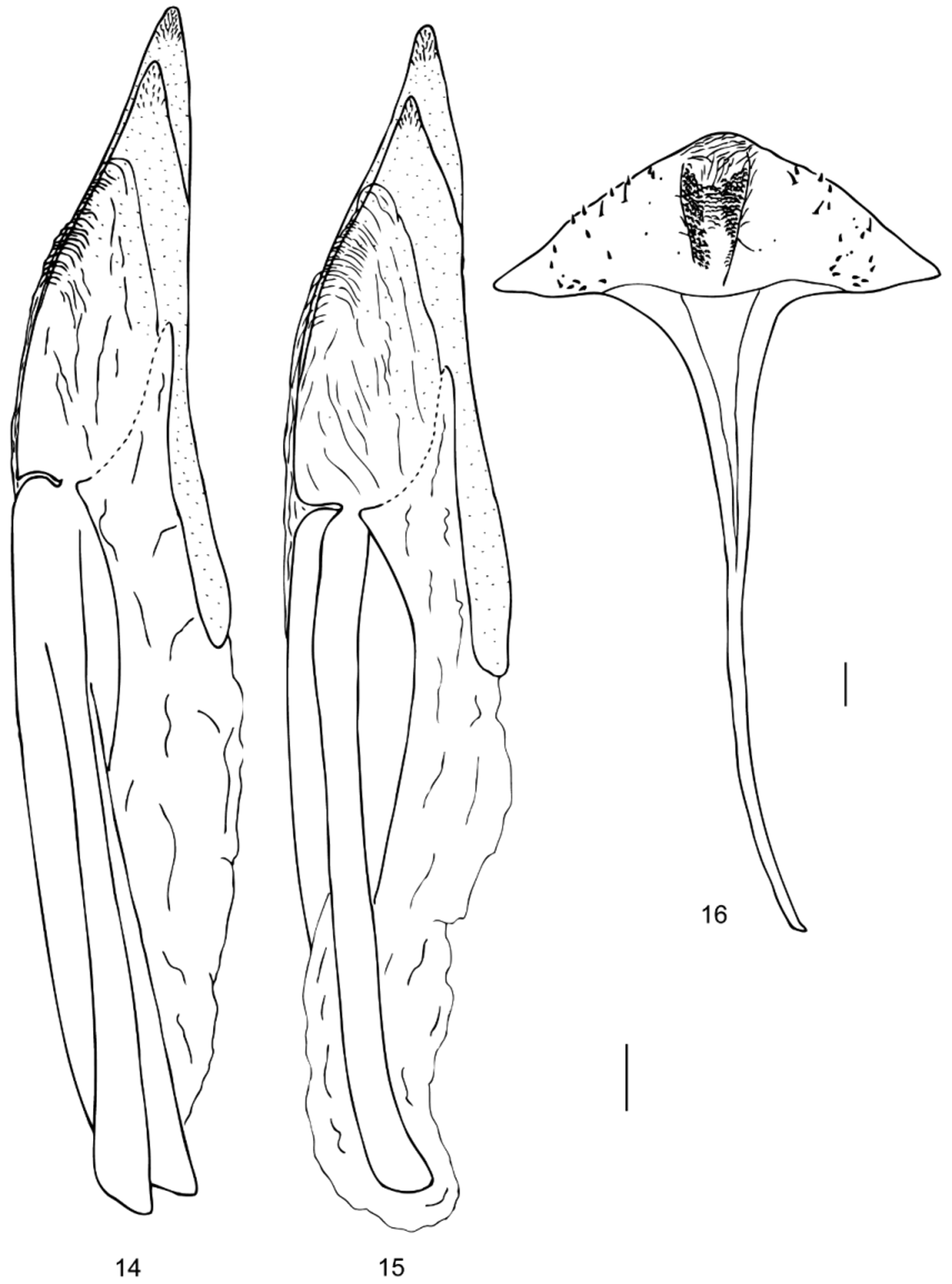
Abdomen with five ventrites, first two connate; ventrite 1 without lateral depression for reception of femora and tibiae; abdominal intercoxal process acute; ventrite 1 longest, ventrite 5 posteriorly slightly excised in males and rounded in females. Male segment VIII: sternite gently rounded posteriorly, not emarginated, distinctly shorter than ventrite 5, with short median process anteriorly ( Fig. 6 View FIGURES 3 – 13 ); tergite more strongly sclerotized laterally than mesally, rounded posteriorly. Female segment VIII: sternite posteriorly sub-triangular, sclerotized laterally, with anterior median process approximately half as long as abdomen ( Fig 16 View FIGURES 14 – 16 ); tergite similar to that of male. Male sternite IX ( Fig. 7 View FIGURES 3 – 13 ) with anterior median process narrow and nearly as long as ventrites 4–5 combined; female sternite IX forming part of ovipositor.
Aedeagus ( Figs 3–5, 8–13 View FIGURES 3 – 13 ) with phallobase robust, long, subcylindrical, moderately curved, 1.14–1.77 (1.40 ± 0.12) times as long as parameres, basally wider than apically (lateral view), with basal orifice asymmetrical, large; parameres individually articulated with phallobase, widest basally, moderately long, gradually curved ventrad and gradually narrowed toward apices; apices rounded in lateral and ventral view. Penis narrow, symmetrical, moderately shorter than parameres, widest basally, gradually narrowed apically; apex narrowly rounded (ventral view); membranous ventral sac with fine longitudinal furrows; fibula narrow, reaching middle of penis (ventral view).
Ovipositor ( Figs 14, 15 View FIGURES 14 – 16 ) about as long as ventrites 3–5 combined, strongly sclerotized; composed of two proximal valvifers and two distal coxites; valvifers wide, long and flattened, medially fused; coxites flattened, plate-like, vertical; left and right coxite slightly asymmetrical, shorter than valvifers, without styli, narrowed posteriorly. Bursa copulatrix tubular, without sclerotized spinules.
Sexual dimorphism. Females are generally larger than males and both sexes differ in the shape and excision of the apex of ventrite 5. The apex of ventrite 5 is slightly excised in males ( Fig. 19 View FIGURES 17 – 20 ), while rounded in females ( Fig. 20 View FIGURES 17 – 20 ). Since ranges of morphometric characters overlap between males and females, they cannot be used for sex recognition of its own.
Variability. The PCA resulted from 12-selected metric characters of males demonstrated that all geographical populations are mutually overlapped with each other in the graph (Fig. 33b). Since the metric data overlaped mutually ( Tabs 1 View TABLE 1 and 2 View TABLE 2 ), we consider specimens from different localities to fit within the species variability. Except for variability of metric characters, specimens moderately vary in the curvature of pronotal sides and in the density of granulation.
Distribution. So far known from Vietnam and Laos ( Fig. 32 View FIGURE 32 ). All specimens were collected using light traps. This species probably represents one of the most commonly light collected dryopid in Vietnam and Laos.
Note. The collection of A. Grouvelle (MNHP) contains a single specimen labelled as: “ Helichus Vaulogeri Grouv / Type / Hanoi, Vaulogue”. We consider this specimen to be conspecific with P. granulosus . To date we have not been able to find any published description of H. vaulogeri , and thus we consider this name as unavailable.
| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parahelichus granulosus ( Delève, 1974 )
| Lojková, Soňa, Degma, Peter & Kodada, Ján 2014 |
Helichus granulosus Delève 1974 : 171
| Deleve 1974: 171 |