
Cymbasoma bitumidum, Suárez-Morales, Eduardo & Mckinnon, David, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4102.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9A7BA798-AA7C-4CAA-B42C-1E260CA573E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6091285 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4CA6D-D51B-FFA9-FF12-509393382D23 |
|
treatment provided by |
Plazi |
|
scientific name |
Cymbasoma bitumidum |
| status |
sp. nov. |
Cymbasoma bitumidum sp. nov.
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 )
Material examined. Adult holotype male from Hamelin Pool Channel, Shark Bay (Station 21 of Kimmerer et al. 1985), Western Australia ( 26°19.770’ S, 114°03.354’ E), partially dissected, ethanol-preserved; dissected parts mounted on 5 slides in glycerine, sealed with Entellan®. Date of collection: 17th June 1983. Slides deposited in the collection of WAM, Australia (cat. WAM C61307).
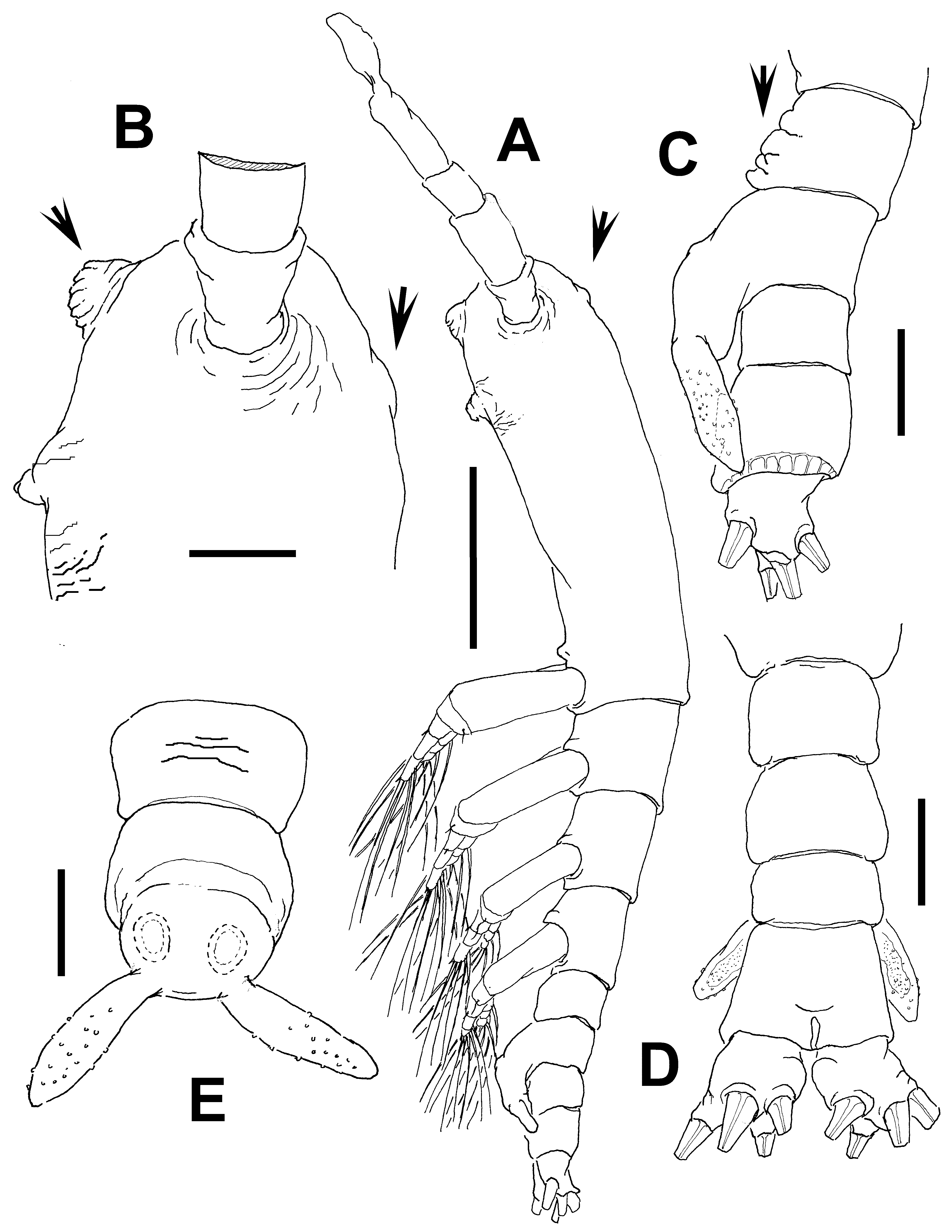
Description of adult male. Total body length 1.2 mm. Cephalothorax 0.57 mm long, representing 50% of total body length ( Fig. 1 View FIGURE 1 A). Midventral oral papilla moderately prominent, positioned at 23% of cephalothorax length ( Fig. 1 View FIGURE 1 A, B). Cephalic region slightly protuberant bilaterally in dorsal view. Pair of dorsal ocelli present, weakly developed; pigment cups relatively small. Ocelli separated by the length of two eye diameters, faintly pigmented. Ventral ocellus relatively small, about same size and diameter as eyes, not easily discernible. No sensilla observed between antennulary bases. Antero-ventral surface of head between antennulary bases and oral papilla bearing cuticular medial rounded protuberance ornamented with transverse ridges (arrowed in Fig. 1 View FIGURE 1 B). Dorsal surface of cephalic area with low medial rounded protuberance (arrowed in Fig. 1 View FIGURE 1 B). Other cuticular processes weak, represented by faint striae set at either side of oral papilla.
Urosome consisting of fifth pedigerous somite, genital somite (carrying genital complex), preanal somite, and anal somite. Fifth pedigerous somite with corrugate ventral surface (arrowed in Fig. 1 View FIGURE 1 C). Genital somite as long as fifth pedigerous somite. Genital complex of type II ( Suárez-Morales & McKinnon 2014), represented by pair of strongly divergent, narrow, leaf-like genital lappets ( Fig. 1 View FIGURE 1 E), these being moderately elongated and dorsally directed in lateral view ( Fig. 1 View FIGURE 1 C), reaching beyond midlength of long anal somite. No medial processes present at flat common basal joint of lappets ( Fig. 1 View FIGURE 1 E) but lappets ornamented with scattered cuticular papillae. Anal somite about twice as long as preanal somite in dorsal view, comprising 33% of urosome length; no suture visible on ventral or dorsal surfaces, but with cuticular hyaline frill along distolateral margins ( Fig. 1 View FIGURE 1 C). Caudal rami subquadrate, approximately as long as wide ( Fig. 1 View FIGURE 1 D), about half the length of anal somite. Each ramus with four setae.
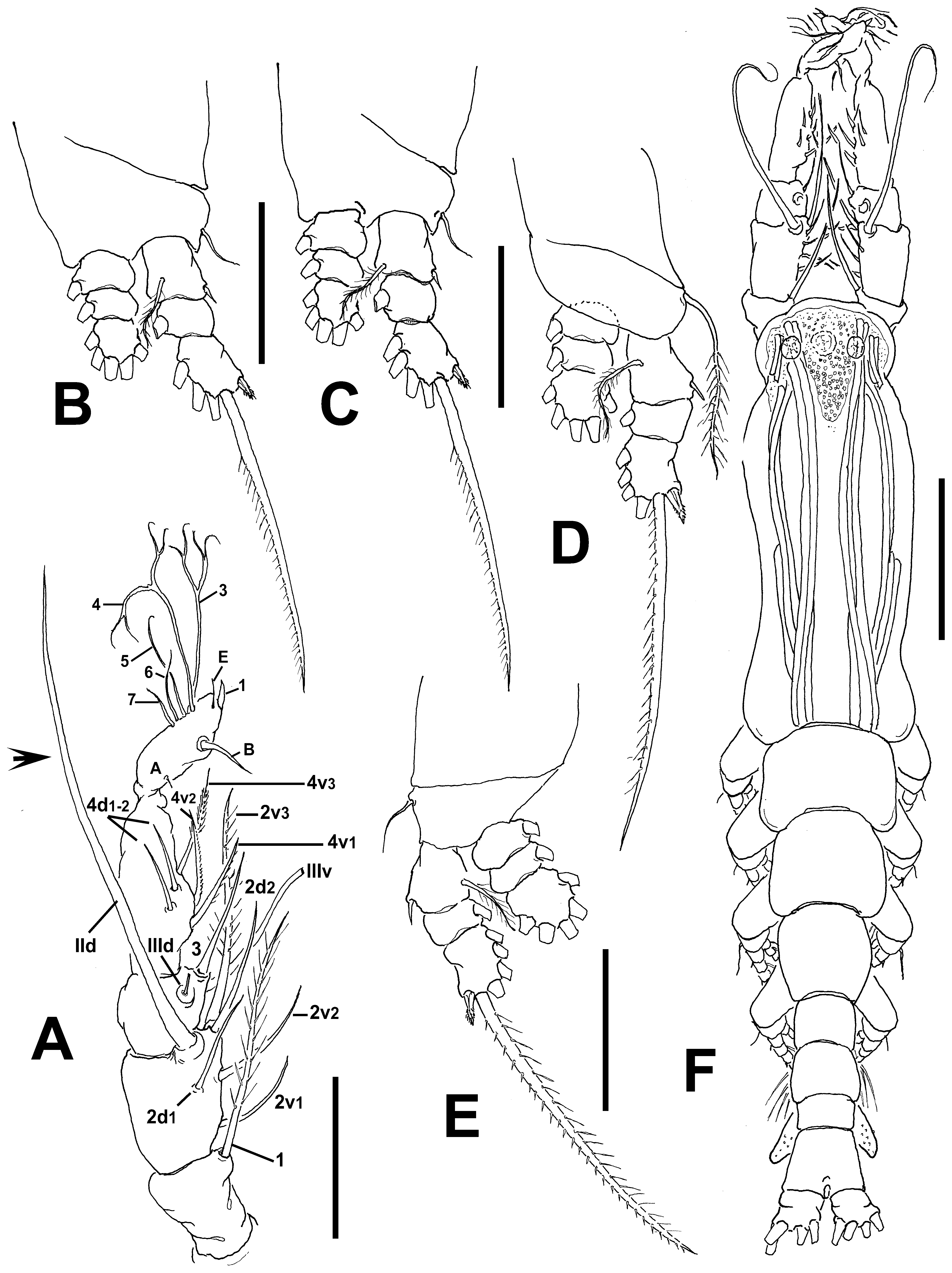
Antennulary length 0.48 mm. Antennules relatively long, representing 40% of total body length, and 80% of cephalothorax length. As usual in male monstrilloids, antennule 5-segmented, all segments separated, with segment 5 located distal to geniculation ( Fig. 2 View FIGURE 2 A). Length ratio of antennulary segments, from first to fifth 16.4: 18.4: 13.3: 26.9: 25 (= 100). Setal element 1 on first segment noticeably long, biserially setulated, reaching proximal 1/3 of fourth segment. Antennulary elements 2v 1-3, 2d1,2, and IId present on second segment, with element 2v 3 noticeably long, setiform, reaching proximal margin of fifth segment and element IId remarkably long, stout, reaching well beyond distal end of fifth segment (arrowed in Fig. 2 View FIGURE 2 A). Elements IIId, IIIv, and 3 present on third segment (element IIId broken off, but socket present). Fourth segment with elements 4d1,2, 4v 1– 3 present; setal elements IVd and IVv absent in specimen. Fifth segment with 5 “b”-group setae, elements b2 and b3 dichotomously branched distally; element 61 present in distal position. According to Huys et al. (2007) setal nomenclature of the distal segment, elements A–E and 1, 2, 6, and 7 present.
Incorporated first pedigerous somite and succeeding three pedigerous somites each bearing well-developed biramous legs. Pedigerous somites 2–4, together accounting for 30% of total body length in dorsal view ( Fig. 2 View FIGURE 2 F). Coxae of each pair unarmed, joined by intercoxal sclerite which is slightly longer than wide. Bases separated from coxae posteriorly by oblique articulation ( Fig. 2 View FIGURE 2 B–E). Exopods of legs 1–4 longer than endopods. Bases of legs 1– 4 with hair-like lateral seta; on leg 3, this seta about 3.2 times longer, sparsely setulated in distal half and slightly thicker than those on the other legs ( Fig. 2 View FIGURE 2 D). Endopods and exopods of legs 1–4 triarticulated. Ramal setae all biserially plumose except spiniform outer seta on exopodal segments 1 and 3. Flexible, slender, sparsely setulated inner seta present on first exopodal segment of legs 1–4. Outer spine on distal exopodal segment of legs 1–4 shorter than segment. Also, outermost apical exopodal setae of legs 1–4 with inner margin sparsely spinulose.
Armature formula of legs as follows:
Female: unknown.
Etymology. The specific epithet is derived from the Latin adjective tumidus (=swollen) preceded by the particle “bi” (=two) to indicate the two protuberances (ventral and dorsal) of the cephalic region.
Type locality. Shark Bay, Western Australia ( 26°19.770’ S, 114°3.354’ E). Hamelin Pool is featured by the presence of stromatolites.
Diagnosis. Cymbasoma with conspicuous corrugate medial protuberance on cephalic area plus low rounded process on dorsal surface of same area. Second antennulary segment bearing extremely long seta 2d2 and stout, long element IId. Antennule representing 40% of total body length and 80% of cephalothorax length. Fifth pedigerous somite with coarsely corrugated ventral surface. Genital complex of type II, with rounded distal tips, smooth basal joint between lappets, ornamented with scattered papillae. Spinous hyaline fringe on lateral surface of anal somite. Four caudal setae.
Remarks. This male specimen was included in the genus Cymbasoma by the presence of two somites between the genital somite and the caudal rami, a key character for males of this genus ( Suárez-Morales & Riccardi 1997; Suárez-Morales 2000a). Only in this genus are males with three caudal setae found, but four caudal setae do occur in some species ( Sars 1921; Suárez-Morales 2000a). This is one of the four species of male Australian Cymbasoma with four caudal setae. Other species sharing this character are: C. quintanarooense ( Suárez-Morales, 1994) , C. tumorifrons (Suárez-Morales, 1999) , C. rigidum Thompson, 1888 , C. longispinosum Bourne, 1890 ( sensu Giesbrecht, 1893), C. bullatum ( Scott, 1909) , C. tropicum ( Wolfenden, 1906) , C. ghardaqanum ( Isaac, 1975) , C. chelemense Suárez-Morales & Escamilla, 1997 , C. rochai Suárez-Morales & Dias, 2001 , and three more Australian congeners described here. This new species can be distinguished from the closely related C. chelemense and its other congeners with four caudal setae by the presence of a conspicuous anterior protuberance with coarsely rugose surface on the cephalic region, which is clearly absent in C. chelemense ( Suárez-Morales & Escamilla 1997: fig. 2B); and by the structure of the genital complex and in the armature of the antennules ( Suárez-Morales & Escamilla 1997: fig. 2C, D). A cephalic protuberance resembling that present in the new species is known also in C. bullatum ( Suárez-Morales 2001c, 2007), but these two species differ in other important characters. In C. bullatum the medial protuberant process is smooth and flanked by two secondary processes ( Suárez-Morales 2007: fig. 2C) whereas the process is corrugated and has no adjacent secondary processes in the new species. In C. bullatum the genital lappets are relatively shorter, not reaching the proximal half of the anal somite (Suárez- Morales 2007: fig. 2F) whereas these lappets almost reach the distal margin of the anal somite in the new species ( Fig. 1 View FIGURE 1 C, D). Also, the medial margin between the lappet bases is clearly protruded and rounded in C. bullatum ( cf. Suárez-Morales 2007: fig. 2E) vs. a flat, smooth margin in the new species. Another species with a strong cephalic protuberance is C. rochai , but in this species the genital lappets are remarkably short, barely reaching the posterior margin of the genital somite, clearly differentiating it from the new species. Also, the medial margin between the lappet bases has a pair of spines in C. rochai ( cf. Suárez-Morales & Dias 2001: fig. 32) which are absent in the new species. Cymbasoma bitumidum has some affinities with the male of C. rigidum , including a flat, smooth medial margin between the lappets and similar body proportions ( Sars 1921), but these species diverge in the structure of the anal somite; in C. rigidum the anal somite has a conspicuous constriction ( Sars 1921) which is absent in the new species. In addition, the distal antennulary segment is differently shaped and has a distinct armature in these species. In C. rigidum the apical element 1 ( sensu Huys et al. 2007) is remarkably long and elements of the “b” group ( sensu Grygier & Ohtsuka 1995) or group 3–7 ( sensu Huys et al. 2007) are short, unbranched, whereas element 1 is short and setae of the “b” group are long and dichotomously branched in the new species. It is also noted that in some of these species with males bearing four caudal setae one of the antennulary elements of the 2v- d group ( sensu Grygier & Ohtsuka 1995) is remarkably long, such as in C. longispinosum ( cf. Giesbrecht 1893), C. tropicum ( cf. Sewell 1949), C. chelemense ( cf. Suárez-Morales & Escamilla 1997), C. rochai ( cf. Suárez-Morales & Dias 2001), C. bullatum ( cf. Suárez-Morales 2007), and also in the new species ( Fig. 1 View FIGURE 1 A). Some males, such as C. bullatum ( cf. Suárez-Morales 2007: fig. 2B), C. rochai ( cf. Suárez-Morales & Dias 2001: fig. 26), and C. chelemense ( cf. Suárez-Morales & Escamilla 1997: fig. 4C, D) also share a plumose, long setal element 1.
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |