
Hyperlasion breviantenna, Sueyoshi & Nakamura & Menzel, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5168.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:AC2A2F68-1EFF-40C2-9886-2268A39EC535 |
|
DOI |
https://doi.org/10.5281/zenodo.6902937 |
|
persistent identifier |
https://treatment.plazi.org/id/C1D159F2-DB03-441A-B2A5-B505E3BF9FE9 |
|
taxon LSID |
lsid:zoobank.org:act:C1D159F2-DB03-441A-B2A5-B505E3BF9FE9 |
|
treatment provided by |
Plazi |
|
scientific name |
Hyperlasion breviantenna |
| status |
sp. nov. |
Hyperlasion breviantenna View in CoL sp. n.
Figs 1 a–f View FIGURE 1 , 2 a–c View FIGURE 2 , 3 a–c View FIGURE 3
Literature. Epidapus sp. — Tagami (2011): 87; Tagami (2014): 20.
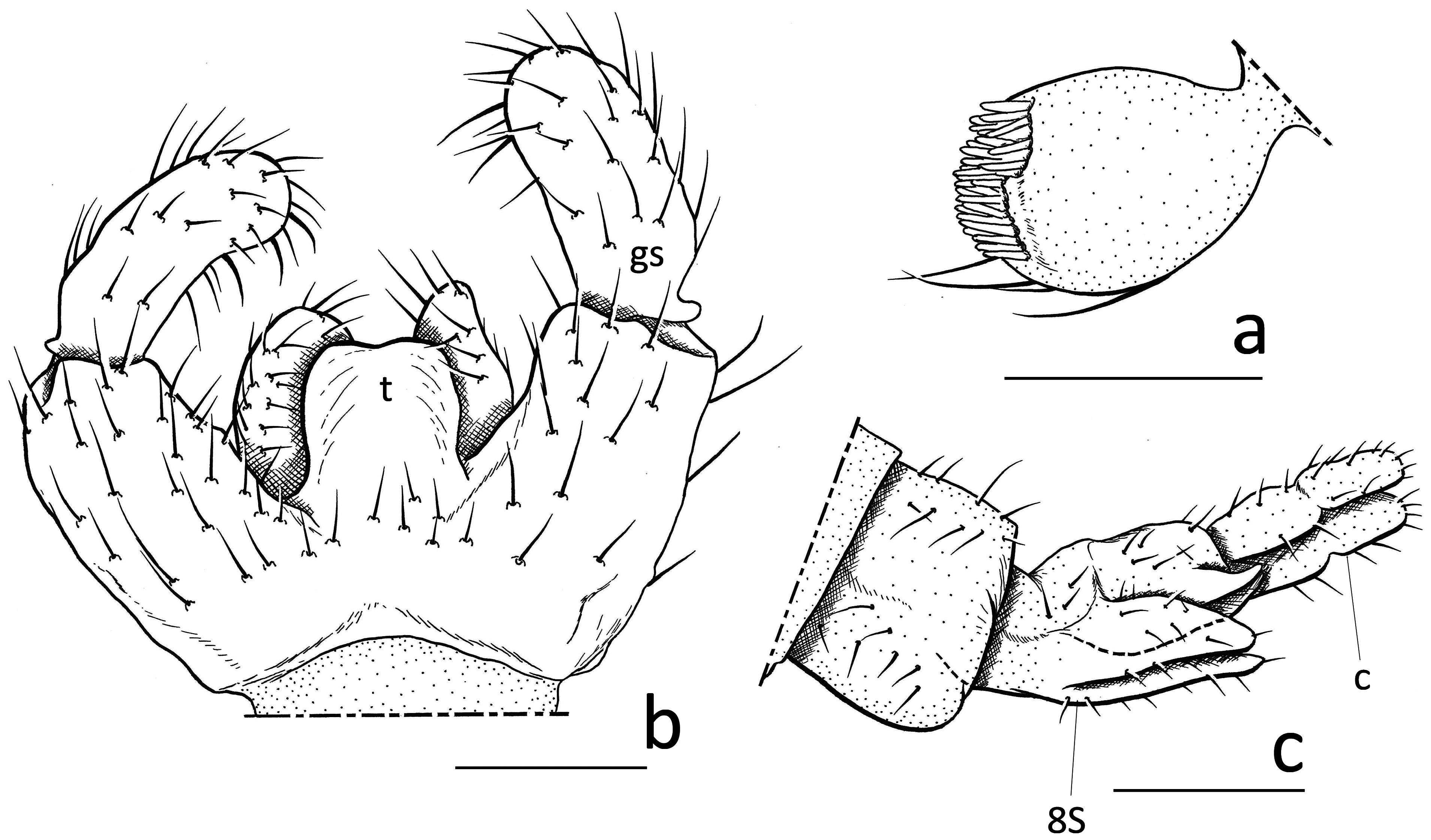
Description. Male. Body ( Fig. 1a View FIGURE 1 ) entirely brown in ground color; all setae concolorous with body sclerites. Body length = 1.3–1.5 mm (n = 5). Head. 3 ocelli present; eye bridge 2 facets wide except 1 facet at middle. Antennae moderately long; scape and pedicel short, roundish and brown (not extended); 4th flagellomere ( Fig. 1c View FIGURE 1 ) 2.0 times as long as wide; basal portion uniform brown, neck paler and hyaline, 1/3 times as long as basal portion of flagellomere; basal portion 1.5 times as long as wide, with coarse setae as long as width of basal part and some long, close-fitting sensilla. Maxillary palpus ( Fig. 2a View FIGURE 2 ) one-segmented with 2–4 ventral setae in equal length; dorsoapically with single large sensory pit and numerous sensilla. Thorax. Pronotum with 4 setae laterally; proepisternum with 5 setae; 2 postpronotal setae present; scutum with 5 presutural and postsutural dorsocentral setae, presutural acrostichal setae; scutellum slightly produced apically, with 6–8 scutellar setae (2 longer setae dorsally and with 2–3 short lateral setae on each side); mediotergite shorter than scutum; katepisternum not extended longitudinally, higher than long. Wing ( Fig. 1d View FIGURE 1 ). Entirely hyaline and slightly tinged with pale brown, 2.6 times as long as wide; posterior wing veins and wing membrane bare; R, R 1 and R 5 only with dorsal macrotrichia; R 1 short, merging with C well before base of M-fork; R 1 = 0.75–0.85 R; stem of M longer than M-fork; M-fork 2.5 times as long as wide; r-m crossvein (y) and stem of M (x) bare; x as long as y; CuA-stem = 1/3 to 1/2 x; c = 0.7 w. Wing length = 1.3–1.4 mm (n = 4). Halter short; stem pale brown, knob brown. Legs. Short and robust, without spines and processes; fore femur thickened, 4 times as long as wide at widest point; fore tibiae shorter than fore tarsus, thickened toward apex, 7.5 times as long as wide at widest point; tibial spurs 1+2+2, short and relatively fine, shorter than diameter of tibiae, in equal length. Claws straight, without teeth. Genitalia ( Fig. 2b View FIGURE 2 ). About as high as wide; gonocoxite short and strong; ventral base of gonocoxite without appendages and processes (without basal lobe or baso-medial groups of setae); tergite X very short and wide, trapezoidal, with 2 rows of coarse setae; tegmen ( Figs 2b View FIGURE 2 : t, 3b) as long as wide, round laterally, with small basal constriction, truncate at apex, without distinct aedeagal teeth; aedeagus ( Fig. 3b View FIGURE 3 ) extremely short, only recognizable as a small drop-shaped structure; gonostylus ( Figs 2b View FIGURE 2 : gs, 3a) about 6/5 times as long as wide, slightly shorter than gonocoxite; apex of gonostylus equally rounded, wider than basal part, covered with short setae, without apical tooth, with 5 megasetae at apical portion; hyaline megasetae hardly distinguishable from other setae on apical portion of gonostylus.
Female. Similar to male but different in the following points. Body ( Fig. 1b View FIGURE 1 ). Entirely brown in ground color. Body length: 1.4–1.8 mm (n = 9). Head. Eye bridge with 2 facets wide, 1 facet at middle. Antennae with globular pedicel and 14 flagellomeres; 4th flagellomere ( Fig. 1e View FIGURE 1 ) as long as wide, neck 1/2 times as long as basal portion; basal portion with more sensilla longer and more strongly curved. Maxillary palpus with 4 or 5 long setae ventrally.
Thorax. Pronotum with 4–5 setae laterally; scutellum reduced as flat sclerite posterior to scutum, apex not produced posteriorly, with long lateral and short apical scutellar setae and adjacent setae around lateral seta; mediotergite enlarged, longer than scutellum, shorter than scutum; proepisternum with more than 10 setae. Wing ( Fig. 1f View FIGURE 1 ). 3.0 times as long as wide; R 1 shorter, = 0.5–0.8 R; c = 0.8 w. Wing length: 1.0– 1.5 mm (n = 13). Legs. Fore tibia 8 times as long as wide at widest point; fore femur 4 times as long as wide. Postabdomen ( Fig. 2c View FIGURE 2 ). S8 divided into longitudinal 2 sclerites; apical segment of cercus ( Fig. 2c View FIGURE 2 : c) slightly shorter than basal segment.
Etymology. The name ‘ breviantenna ’ is Latin ( brevis = short; antenna = antenna). This specific epithet is a noun in apposition that refers to the short female antenna of this species.
Specimens examined. Type material. Holotype: ♂, JAPAN, “Ichinokura, Tajimi City, Gifu Prefecture, Japan, 1.vii.2021, M. Sueyoshi leg.” ( Hb. 0060 in BLKU) . Paratypes: JAPAN. Shizuoka Prefecture: 10♀, Ohya, Shizuoka City , 23–30.vi.2016, Y. Tagami leg., collected by yellow pan traps ( Hb. 0023– Hb. 0034) . Gifu Prefecture: 12♀ , Ichinokura, Tajimi City , 16.vii.2020, Y. Tagami leg. ( Hb. 0045– Hb. 0056) ; 2♂, same data as holotype (Hb.0061, Hb. 0062) ; 1♂, same date and locality as holotype, Y.Tagami leg. ( Hb. 0059) ; 15♀, Kasahara, Tajimi City , 17.vii.2020, anonymous collectors ( Hb. 0001– Hb. 0015) ; 3♀, Asahigaoka, Tajimi City , 17.vii.2020, anonymous collectors ( Hb. 0020– Hb. 0022) ; 1♂ 1♀, Nisizaka, Tajimi City , 18.vii.2020, anonymous collectors ( Hb. 0016, Hb. 0017) . 1♂ 11♀, Fukuoka Prefecture: Dazaifu City , 10.vi.2016, K. Kawagoe leg. ( Hb. 0033– Hb. 0044) .
Other specimens (non-type series). JAPAN (all preserved in ethanol and preserved in FFPRI): Gifu Prefecture: 869♀, Kasahara, Tajimi City , 17.vii.2020, anonymous collectors ; 32♀, Shiominomori, Tajimi City , 25.vi.2021, Y. Tagami leg. ; 996♀, same locality and date, M. Sueyoshi leg. ; 23♀, Higashiyama, Tajimi City , 1.vii.2021, M. Sueyoshi leg. ; 48♀, Sannokura, Tajimi City , 1.vii.2021, M. Sueyoshi leg. AUSTRALIA ( BIN BOLD: ACQ9549 ; all data taken from BOLD). New South Wales: 2♀, Glendale Road, Berowra, Brooklyn [= Glendale Road, NE of Berowra, N of Sydney], - 33.5990°N, 151.1640°E, 205m, Malaise trap, 27.xii.2012, P. Hebert leg. ( BIOUG15885 View Materials - H03, BIOUG15886 View Materials -G04). GoogleMaps 2♀, Hat Head [= Hat Head, S of South West Rocks, S of Coffs Harbour], - 31.0626°N, 153.0520°E, 36m, 18.ii.2012, P. Hebert leg. ( BIOUG37606 View Materials -A05, BIOUG37606 View Materials -F02) GoogleMaps .
Specimens used for molecular analysis (not preserved as physical vouchers). JAPAN. Gifu Prefecture: 6♀, Kasahara, Tajimi City , 17.vii.2020, anonymous collectors ( Hyperlasion-breviantenna _female_JPN-Gifu_ 0607 T10 , Hyperlasion-breviantenna _female_JPN-Gifu_ T1 to Hyperlasion-breviantenna _female_JPN-Gifu_ T6 ) ; 1♂, Ichinokura, Tajimi City , 1.vii.2021, Y. Tagami leg. ( Hyperlasion-breviantenna _male_JPN- Gifu _202107) ; 1♀, Ohya, Shizuoka City , 23–30.vi.2016, Y. Tagami leg., collected by pan traps ( Hyperlasion-breviantenna _female_ JPN-Shizuoka_S4) .
Distribution. Japan (Honshu, Kyushu). Australia ( New South Wales).
Remarks. Hyperlasion is characterized in Menzel & Mohrig (2000) by the following morphological characters: maxillary palpus one-segmented, with sensory pit apically; wing vein R 1 shorter than R, merging with the costal vein C before base of M-fork ( Fig. 1d View FIGURE 1 ); anal area of wing reduced ( Fig. 1d View FIGURE 1 ); halter with short stem; apex of fore tibia undifferentiated, without distinct group of setae; apex of gonostylus rounded, toothless and with mesal megasetae only at the inner side of the gonostylus ( Figs 2b View FIGURE 2 , 3a View FIGURE 3 ). Our new species is distinguished from other Hyperlasion congeners by the very short female antenna ( Fig. 1b View FIGURE 1 ) with flagellomeres as long as wide ( Fig. 1e View FIGURE 1 ), the gonostylus with more than four (mostly 6 to 7) mesal megasetae ( Fig. 3a View FIGURE 3 ), and a membranous, almost quadrangular tegmen longer than wide ( Fig. 3b View FIGURE 3 ), similar to H. wasmanni . Hyperlasion breviantenna sp. n. differs from this species by the much shorter female flagellomeres, the thickened maxillary palpus with a larger sensory pit in both sexes, slightly longer gonostyli with an evenly rounded apex, much weaker mesal gonostylar megasetae, and an extremely short aedeagus ( Fig. 3b View FIGURE 3 ).
Sexual dimorphism (except for the genitalia) of this species is observed in: male antennal flagellomeres longer than those of female; and fore tibia of male thicker than that of female. Other morphological differences between males and females are not observed in wing shape, height/length of the katepisternum, and leg length such as reported in H. aliens ( Menzel & Smith 2009) and H. viridiventris ( Menzel et al. 2013) .
Tagami (2011, 2014) gave it the Japanese name “Shizuoka-kohige-kurobane-kinokobae”. We assume that it includes most of black fungus gnats which have been called locally as Japanese “kobae” [= gnats] ( Tajimi City 2020), “tajimimushi” [= gnats of Tajimi City], and “kurobane-kinokobae” [= sciarids] ( Goto 2014) in Tajimi City.
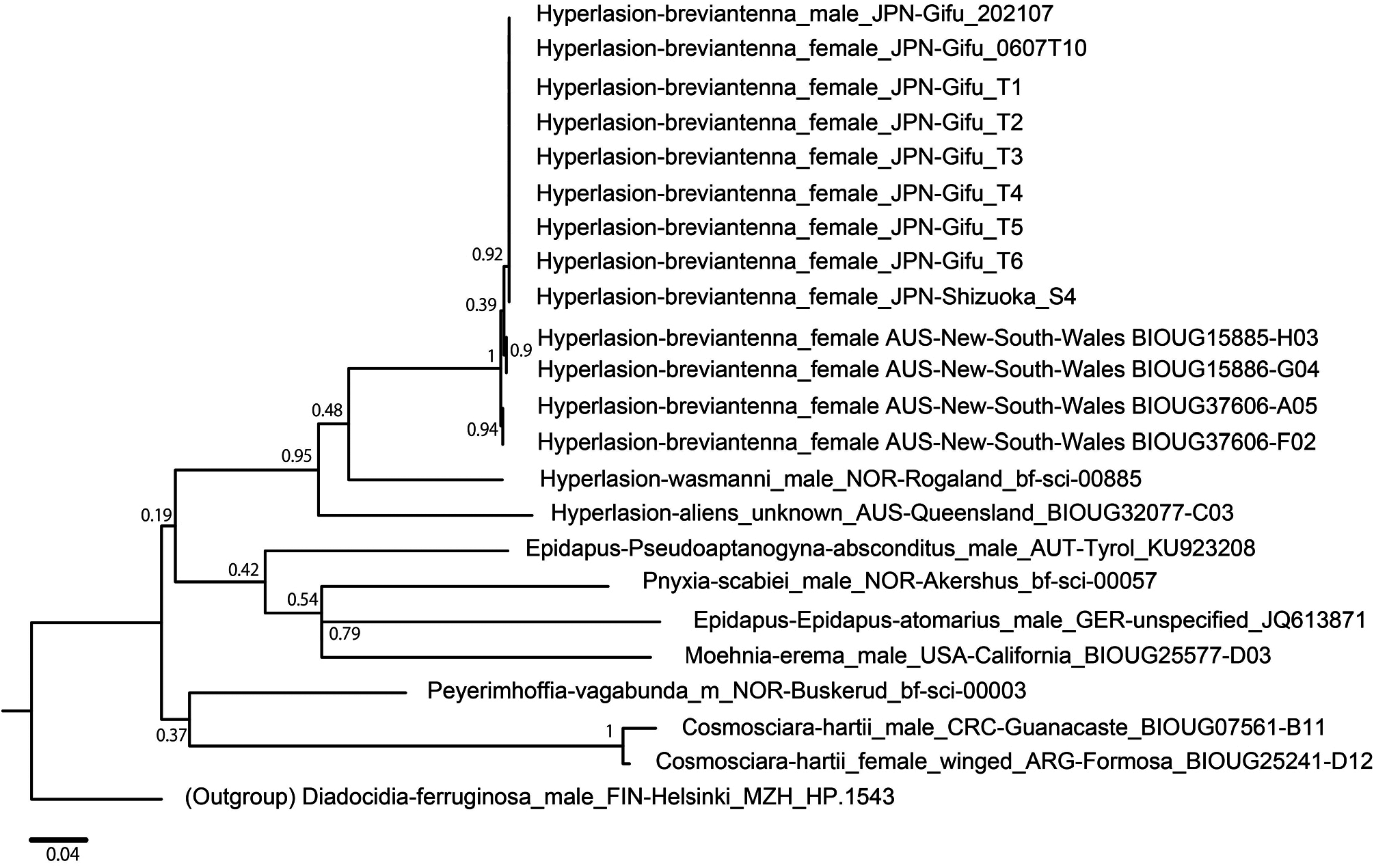
Taxonomy based on genetic analysis. A ML tree rooted with Diadocidia ferruginea as an outgroup of sciarids is shown in Fig. 4 View FIGURE 4 . It supported the monophyly of the Hyperlasion species examined here. It further indicates that the Japanese and Australian specimens are clustered as in a one monophyletic group with less than 1% differences in the examined nucleotide sequences, and it was distinctly different from other congeners examined. Thus, we can expect that these specimens are from a distinct species of Hyperlasion , which we refer to here as H. breviantenna . The Australian females are conspecific with Japanese specimens due to the small genetic distance of less than 1 % to the Japanese specimens ( Fig. 4 View FIGURE 4 ). Australian records lack any male specimens. No males are known from Australia so far.
Biology of Hyperlasion breviantenna in Japan
This species has been reported as a nuisance pest, after vast number of adults infested in urban areas of Tajimi City, Gifu Prefecture, Japan ( Goto 2014) and Shizuoka City, Shizuoka Prefecture, Japan ( Tagami 2014). In urban areas, the flies enter houses through gaps between closed windows. The adults emerge early in the morning on sunny days during the rainy season, i.e. June to July ( Tagami 2011, 2014). More than 100 individuals enter rooms per half hour, peaking at around 09:00–10:00 and declining at noon ( Tagami 2011, 2014). Tagami (2011) reported that most of the flies invading houses were females. After entering the buildings, they soon die ( Fig. 3a View FIGURE 3 ). The adults which were mentioned in the literature were found at the forest edge ( Tagami 2014), on humus ( Goto 2014) and mown grass ( Tajimi City 2020) in Japan.
The plagues of H. breviantenna in Japan are obviously female-biased. The specimens collected at three localities in Tajimi City on 1 July 2021, include just three males but more than a thousand females simultaneously. We found two previous reports on sciarid mass occurrence in the literature. Pritchard (1960) reported that thousands of females of Moehnia erema occurred in the Entomological Laboratory of the University of California, USA, during three days in August. Heller et al. (2009) reported 2618 females of H. wasmanni with malaise traps in three weeks during July and August, 2004, in Sweden. Nielsen et al. (2016) reviewed published collection data of H. wasmanni and noted that the occurrence of the species is skewed towards female. Steffan (1981) stated that M. erema is parthenogenetic, with few male occurrences. We found a small number of males in the field. This implies that H. breviantenna must not parthenogenetic, but we have no data to substantiate this at the present time.
Hyperlasion breviantenna is assumed to inhabit forests and forest edges ( Tagami 2014; Goto 2014) and further details are unknown at present time. Only of the most common Hyperlasion , the widespread type species H. wasmanni Schmitz , are the ecology and lifestyle better studied, including habitats. Hyperlasion wasmanni is common in various vegetation: woodlands, heathland, sedge beds, raised bogs, and damp meadows (e.g., Menzel et al. 2006). Schmitz (1918) and de Meijere (1928) reported that H. wasmanni was collected from nests of ants: Lasius fuliginosus (Latreille) , L. niger (Linnaeus) , and Formica exsecta Nylander ( Hymenoptera : Formicidae ), in the Netherlands. Heller et al. (2009) and Nielsen et al. (2016) referred to it as myrmecophilous. Steffan (1974) reported that larvae of H. wasmanni (as a junior synonym, H. magnisensoria Hardy, 1956 , in the text) were found on fallen Coprosoma logs covered with moss, and that they fed off the surface of the wood under the thick mat of moss. Extensive studies to microhabitats in which the development cycle of H. breviantenna takes place and the host plants of or the fungi associated with H. breviantenna larvae still need to be carried out. Further researches focusing on any ants and mosses when seeking habitats and biological interactions of H. breviantenna to other organisms might help in the development of control measures for this new pest insect.
H y perlasion breviantenna may have genetically closed populations outside of Japan and Australia. Ten sciarid species are known as cosmopolitan pests in agriculture ( Broadley et al. 2018). Sciarids are easily transported with plants and soil. We suspect that one or both populations may have been introduced via cultivated plants and have become established in Japan and/or Australia. Some literature reports natural dispersal with long distances between the mainland and oceanic islands or between them have been reported ( Holzapfel & Harrell 1968; Hardy & Cheng 1986; Peck 1994). We need sufficient evidence on natural and artificial events associated with dispersal of Hyperlasion breviantenna .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |