
Bairdia jeromei, Forel, 2012
|
publication ID |
https://doi.org/ 10.5852/ejt.2012.19 |
|
publication LSID |
lsid:zoobank.org:pub:893DB151-7B86-485C-932F-A01C57B20316 |
|
DOI |
https://doi.org/10.5281/zenodo.3858794 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB2C0F-FF8E-4E1A-D408-F31AFE5E3312 |
|
treatment provided by |
Carolina |
|
scientific name |
Bairdia jeromei |
| status |
|
Callicythere postiangusta Wei, 1981
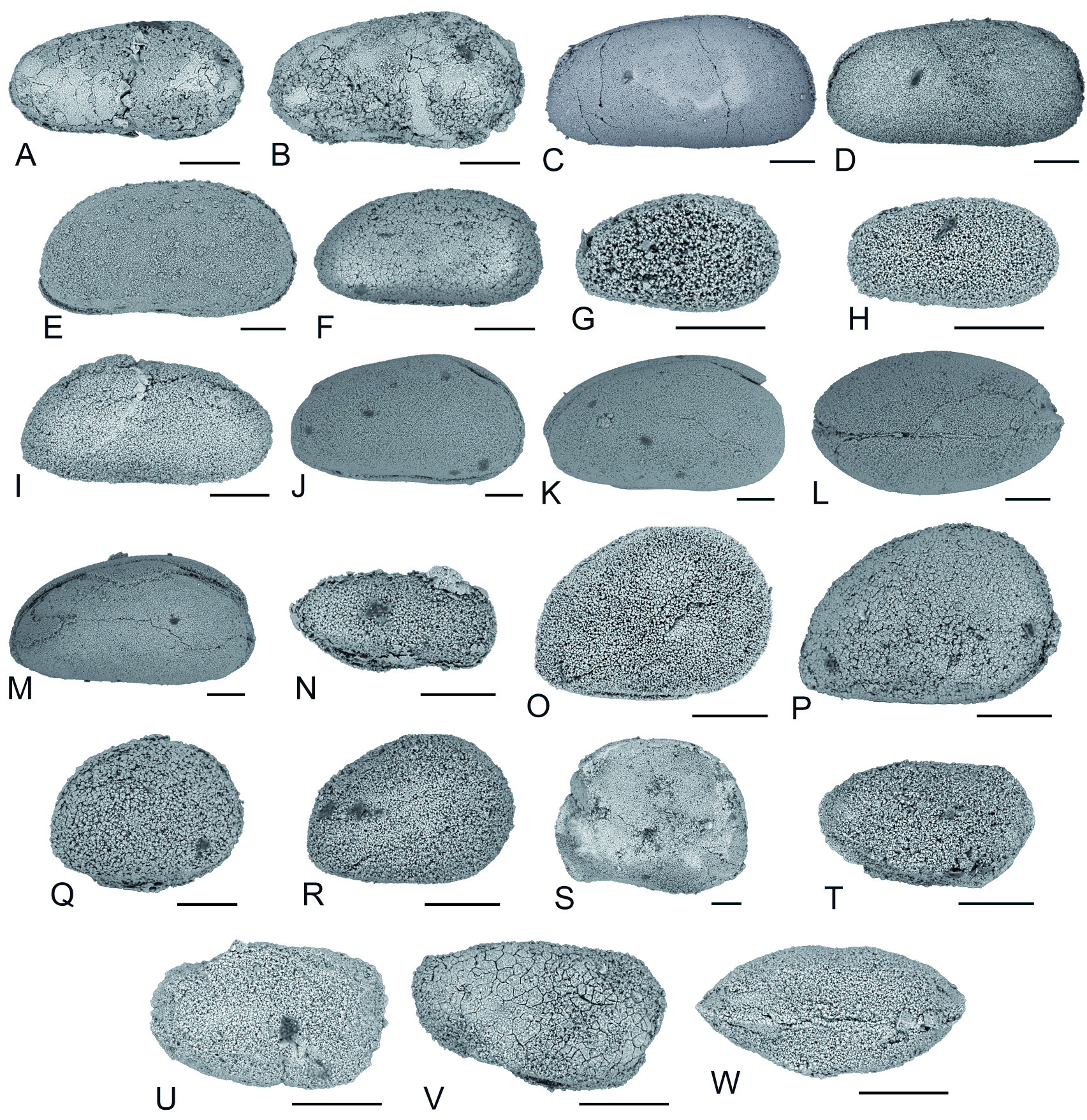
Figs 14 View Fig T-W, 15A-B
Callicythere postiangusta Wei, 1981: 504, pl. 1, figs 19-22.
Callicythere postiangusta – Crasquin-Soleau & Kershaw 2005: pl. 1, figs 1-6.
Localities
- Samples 05PAJ30, 31, 33,35,37, 38,44,Daye Formation, Dajiang section(25°33’56’’N- 106°39’41’’E),
Guizhou Province, South China, Griesbachian, Early Triassic. - Weiyuan, Leikoupo Formation, Sichuan Province, South China, Middle Triassic ( Wei, 1981). - Çürük dağ section, Western Taurus, Turkey, Early Triassic (Forel, in progress). - Laolongdong section, Feixianguan Formation, Sichuan Province, South China, Early Triassic
( Crasquin-Soleau & Kershaw, 2005).
Ostracod biodiversity variations
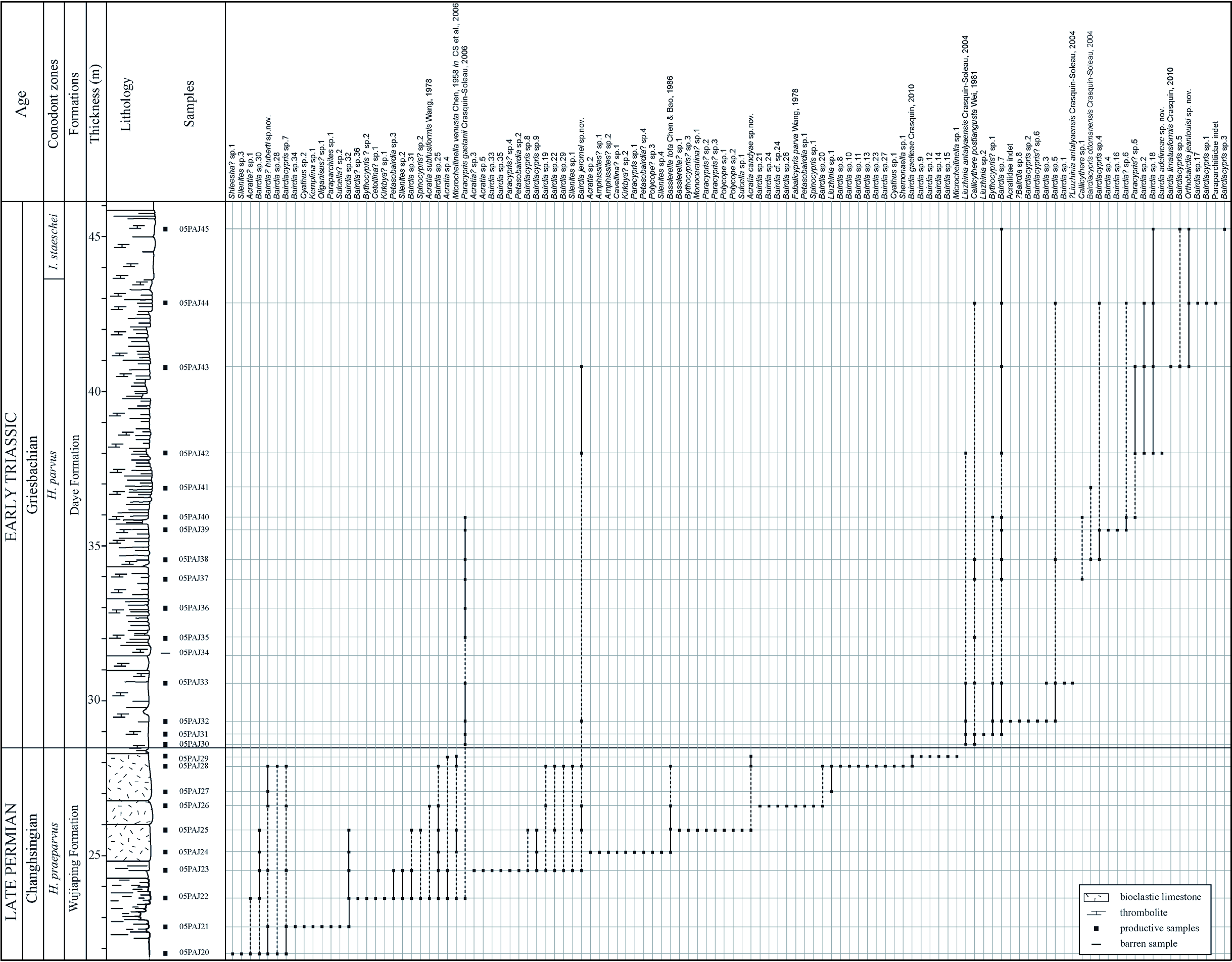
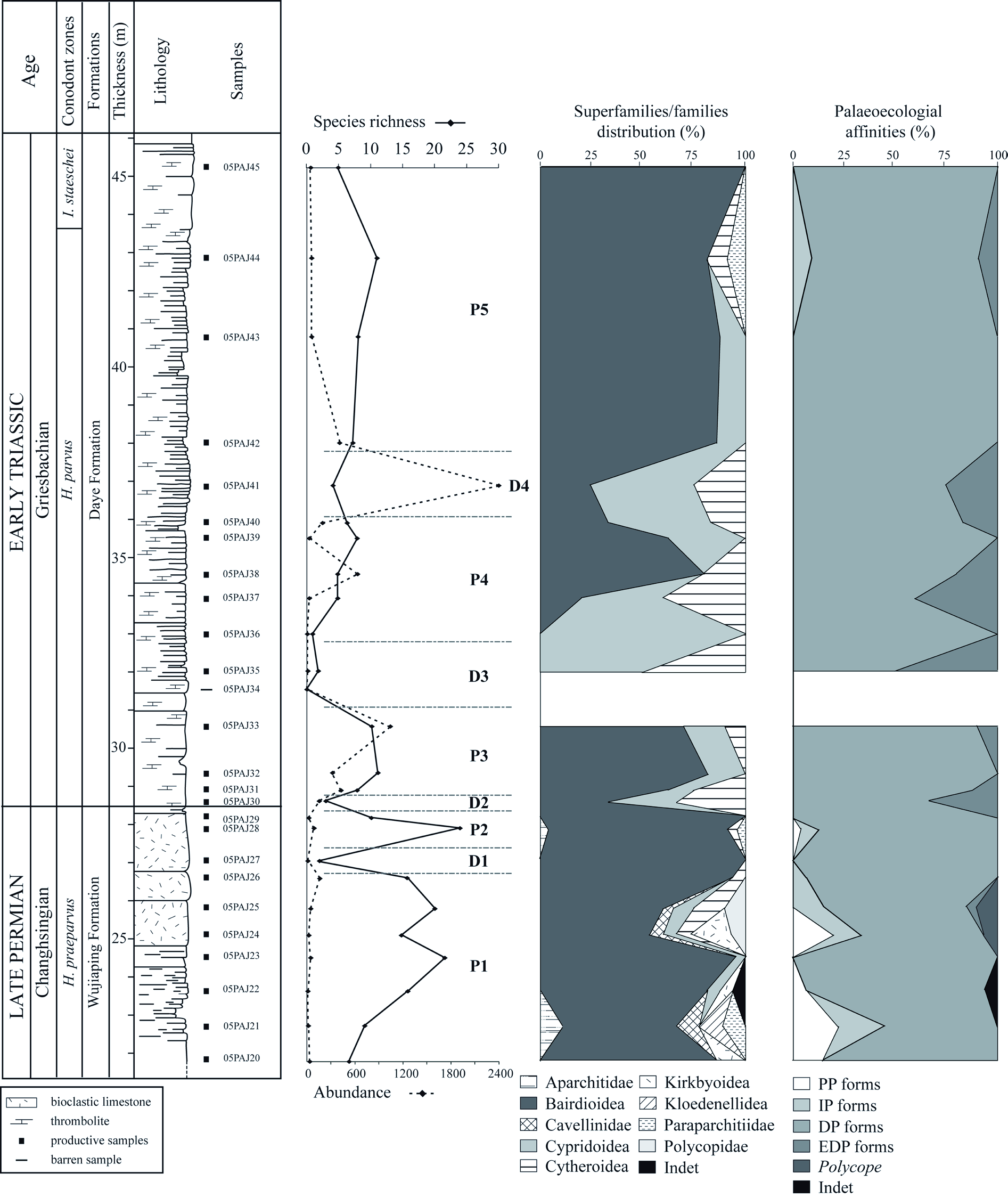
Ostracods are found from the top of the Wujiaping formation (05PAJ20) to the base of the Daye formation (05PAJ45). Of the 26 samples within this interval, 25 yielded ostracods. Species distribution is summarised in Fig. 2 View Fig and abundance and species richness variations are presented in Fig. 16A View Fig . In the productive samples, abundance varies from 1 (05PAJ36) to 2403 (05PAJ41) and species richness from 1 (05PAJ36) to 24 (05PAJ28). A taxa-specimen rarefaction analysis has been performed and suggests that communities have been well sampled and little sampling bias is expected. Several peaks (P) and drops (D) are distinguished on the biodiversity curves ( Fig. 16A View Fig ):
(1) Samples 05PAJ20 to 26 are relatively highly diversified, for both species richness and abundance (P1).
(2) This phase is followed by a sharp drop from 20 species in 05PAJ25 to 2 in 05PAJ27 (D1). 05PAJ27 records the minimum of Permian ostracod diversity in Dajiang.
(3) An important diversification is observed just before the Permian – Triassic boundary (PTB) (P2: 05PAJ28, 29). This peak is the maximum of species richness in Dajiang.
(4) Samples 05PAJ28 to 05PAJ30 (D2) bracket the PTB and show an important reduction of specific richness (from 24 to 3).
(5) Abundance and species richness then increase from 05PAJ31 to 05PAJ33 (P3). Species richness is however lower than in the Permian assemblages.
(6) Another reduction of diversity, both abundance and specific richness, is recorded in 05PAJ34-36 (D3).
(7) A slight rediversification is observed from 05PAJ37 to 05PAJ40 (P4).
(8) A reduction of species richness, together with a high abundance, is recorded in 05PAJ41 (D4).
(9) 05PAJ42 to 05PAJ45 (P5) show a slight rediversification of the assemblages. During this phase, species richness is relatively stable while abundance is low.
The biodiversity parameters in Dajiang show striking inversion through the PTB: high species richness/ low abundance for Permian assemblages, low species richness/ high to very high abundance in Triassic microbialites.
Composition of assemblages
Ostracods in Dajiang belong to 10 superfamilies/families, the distribution of which through the PTB is illustrated in Fig. 16B View Fig . The most abundant superfamily, both in the Permian and the Triassic, is Bairdioidea (encompassing the genera Acratia , Bairdia , Bairdiacypris , Bythocypris , Fabalicypris , Kempfina , Liuzhinia , Microcheilinella , Orthobairdia, Petasobairdia , Silenites and Spinocypris ). It is absent from only 2 assemblages from D3 (05PAJ35, 36). Its proportions are between 20% (05PAJ37) and 100% of species (05PAJ27, 29, 45). The second most important superfamily is Cypridoidea (genus Paracypris ). It is ubiquitous both in Permian and Triassic assemblages but shows higher proportions in Triassic assemblages. It is absent from 05PAJ20, 21, 26-29, 38, 44, 45. When it is present, it represents between 6% (05PAJ22) and 100% (05PAJ36) of the species. Cytheroidea (genera Basslerella, Callicythere and Monoceratina ) are found both in Permian and Triassic assemblages. When it is present, it represents between 4% (05PAJ28) and 50% (05PAJ30, 35) of species. Paraparchitidae (genera Paraparchites, Shemonaella, Paraparchitiidae indet.) are present in 2 Permian assemblages and 1 Triassic: 05PAJ21 (11%), 05PAJ28 (4%), 05PAJ44 (9%). The following superfamilies/families only occur in the Permian: Cavellinidae (genera Cavellina , Sulcella ) are found in only 3 assemblages of P1 (05PAJ21, 24, 25) where they represent between 5% (05PAJ25) and 11% of species (05PAJ21). Kirkbyoidea (genera Amphissites, Kirkbya, Shleesha) is part of 3 assemblages from P1 (05PAJ20, 22, 24) where it is respectively 14, 6 and 20% of the species. Polycopidae (genus Polycope ) is present in 2 assemblages from P1: 05PAJ24 (7%) and 05PAJ25 (10%). Kloedenellidea (genus Oliganisus ) is found in only one assemblage (05PAJ21) where it represents 11% of the species. Aparchitidae (genus Cyathus ) is present in 2 assemblages: 05PAJ21 (11%) and 05PAJ28 (4%). Finally, only one species belongs to an undetermined superfamily/ family (genus Cetollina ). It is part of 05PAJ22 where it represents 6% of species.
In terms of species, the main extinction event occurs in the latest levels of the Wujiaping formation and the extinction rate is of 98%. Two species cross the PTB in Dajiang: Bairdia jeromei sp. nov. and Paracypris gaetanii Crasquin-Soleau, 2006 ( Fig. 2 View Fig ). They are found in almost all microbialites, their last occurrences are high in the section (respectively 05PAJ41 and 05PAJ43). Two of the ten genera found in the microbialites have not been recorded in Permian strata before: Callicythere and Orthobairdia . Consequently, the faunal turnover is nearly complete at the specific level (94% of new species), partial at the generic level (20% of new genera) and no renewal is observed at the superfamily/family level. Importantly, most assemblages from the microbialites show high proportions of very small specimens (100 μm in length). These specimens belong to the same genera as the larger forms: hence the deviation of size is not due to the presence of intrinsically smaller genera. This aspect of the faunas will be addressed in future work.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Cytheracea |
|
Family |
|
|
Genus |
Bairdia jeromei
| Forel, Marie-Béatrice 2012 |
Callicythere postiangusta
| Callicythere postiangusta Wei, 1981: 504 |
Callicythere postiangusta
| Crasquin-Soleau & Kershaw 2005 |