
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/10.1073/pnas.1216534110 |
|
DOI |
https://doi.org/10.5281/zenodo.3809064 |
|
persistent identifier |
https://treatment.plazi.org/id/03B887AE-A72C-F054-FF88-FC4CBFD1FC86 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
Two coalesced hadrosaur (compare with Edmontosaurus annectens) caudal vertebrae were discovered in the Hell Creek Formation of Harding County, South Dakota ( 40). Archosaur fauna identified in this site include crocodiles, dinosaurs, and birds ( 41). Physical evidence of dental penetration and extensive infection (osteomylitis) of the fused vertebral centra and healing (bone overgrowth) document an unsuccessful attack by a large predator.
A tooth crown was discovered within the wound, permitting identification of the predator as T. rex .
This is unambiguous evidence that T. rex was an active predator, fulfilling the criteria that Farlow and Holtz ( 3) advanced. As T. rex comprises between 1% and 16% of the Upper Cretaceous dinosaurian fauna in Western North America ( 41 – 45), its status as a predator or obligate scavenger is nontrivial and could have significant implications for paleoecological reconstructions of that time period. The present contribution provides unique information demonstrating the ecological role for T. rex as that of an active predator. Despite this documentation of predatory behavior by T. rex , we do not make the argument that T. rex was an obligate predator. Like most modern large predators ( 27, 45) it almost certainly did also scavenge carcasses ( 9, 16).
Results
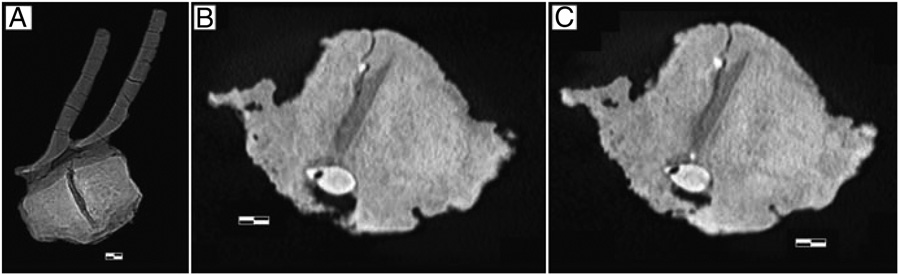
The fused vertebrae are an articulated pair from the midcaudal series, estimated to have been in positions between caudals 24 and 31, of an adult hadrosaur, most likely E. annectens ( Fig. 1 A and B View Fig. 1 ). The dimensions of both vertebrae are nearly identical, with a dorsal process 15.5 cm in length and a centrum length of 5.5 cm. Although the specimen shows some abrasion caused by fluvial transport, there does not appear to have been an appreciable loss of exterior bone and it is well preserved overall. Fusion of the vertebrae was accomplished through extensive rugose syndesmophyte-like bone growth that covers the majority of the external surface of the centra ( Fig. 2 A–C View Fig. 2 ). This reactive bone massively overgrew the entire external surface of the two vertebrae. The articular surfaces of the centra appear to have been spared and are completely free of reactive bone. Postmortem breakage on the ventral surface of the centra indicates that the chevron could also have been fused to the centra, but had been subsequently broken off during fluvial transport. A tooth of a large theropod was situated ventrolaterally on the left side, lodged between the two vertebral centra. The tooth consists of the crown only, with no root material present. The rugose bone growth on the centra extends up to and around the embedded tooth, partly enclosing it ( Figs. 1 View Fig. 1 and 2 View Fig. 2 ).
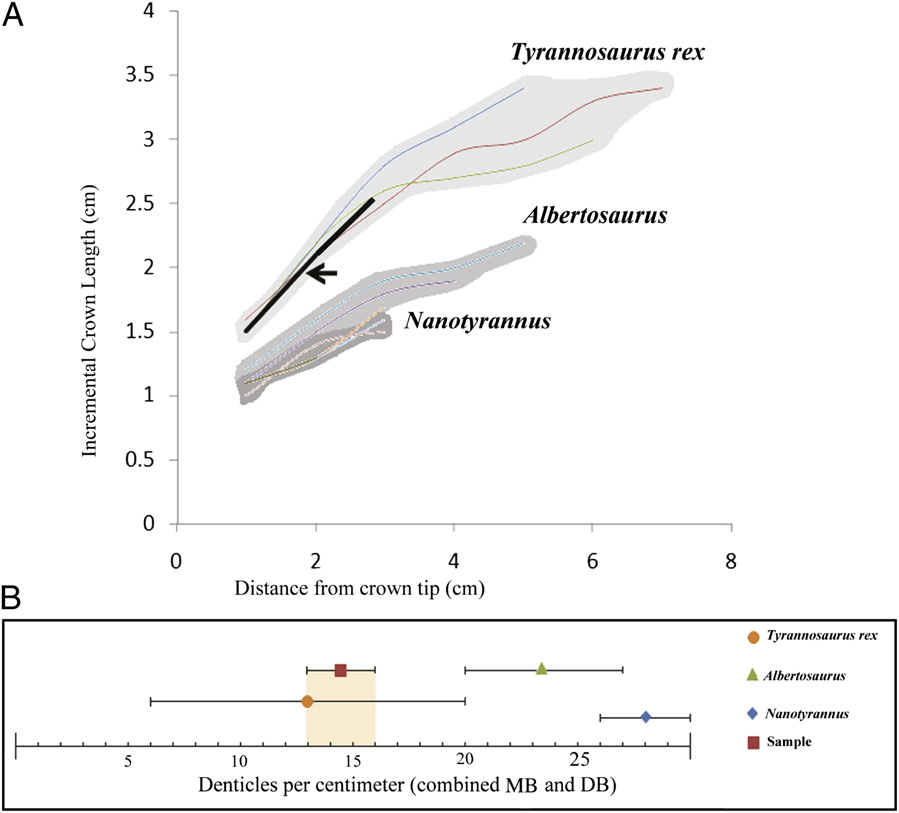
The theropod tooth crown is well preserved, with the broken basal portion of the tooth partially exposed ( Fig. 1 View Fig. 1 ). Computerized tomographic (CT) scans revealed the crown height to be 3.75 cm, and visual inspection revealed the crown base length to be 2.35 cm and the crown base width 1.65 cm. Denticles are well preserved on the mesial and distal carinae ( Fig. 3 View Fig. 3 ), and the distal basal denticle (DB) and mesial basal denticle (MB) densities are 16 per cm and 13 per cm, respectively. Comparison of the embedded tooth’ s dimensions and morphometric relationships with the data from the Smith et al. ( 46) study reveals a strong alliance with T. rex ( Fig. 4 View Fig. 4 ). The tooth is indistinguishable in morphology, size, and denticle character from known T. rex subadults (e.g., Los Angeles County Museum–23845 View Materials and Black Hills Institute–6439 ). An independent comparison of the ratio of the distance from crown tip (DCT) to the incremental crown length (ICL) for T. rex and Nanotyrannus , the only two contemporaneous large-bodied and large-toothed theropods ( 46 – 49), with that of the embedded tooth places it firmly within the T. rex range ( Fig. 3 View Fig. 3 ). For this study, Albertosaurus was added as a control. In addition, study of the embedded tooth’ s denticle density indicates that its DB and MB values overlap those of only one animal studied, T. rex ( Fig. 3 View Fig. 3 ). Morphologic and morphometric characters of Nanotyrannus are sufficiently dissimilar from the embedded tooth to exclude it from candidacy for the tooth-producing taxon. Only one animal studied— T. rex —bears close resemblance to the tooth in question.
Discussion
The rugose structures observed on the hadrosaur vertebrae are consistent in morphology with bone growth associated with healing injuries, as observed in modern and extinct animals ( Fig. 1 View Fig. 1 ). In modern endothermic animals, trauma to bone is followed by signs of insipient bone healing within the first several weeks. The first macroscopic evidence of bone healing in mammals can be recognized 7–10 d after the injury ( 50). Healing of reptilian bone is much slower ( 50) and takes longer in reptiles ( 50 – 53), significantly delaying opportunity for its recognition on macroscopic examination. The massive bone reaction in this report suggests survival of the injury for a significant period, perhaps even years. The injury does not appear to have contributed to the demise of this hadrosaur.
The usefulness of individual theropod teeth in determining their taxonomic affinities has been recognized by other researchers ( 46, 54) and detailed, quantitative methods of examining theropod teeth have recently been developed and refined ( 46). The methods developed by Smith et al. ( 46) demonstrate the extent of the discriminating potential of theropod dental characters and enabled identification of isolated T. rex tooth crowns with over 96% accuracy in their study.
The localization of the tyrannosaur tooth in the midcaudal area of the hadrosaur is consistent with that noted in modern pursuit predator attacks ( 27, 55). Kalahari lions have been observed to initially target the hindquarters of the prey animal, in an attempt to immobilize it. This hadrosaur escaped and began to heal around the tooth lodged in its caudal vertebrae, providing evidence of the attack and the identity of the attacker. Such evidence is rare in the fossil record for good reason—prey rarely escapes. As such, it is plausible that many of the purported T. rex bite marks that show no healed bone are actually indicative of successful kills. What is clear is that the T. rex did not finish off this hadrosaur.
The fused vertebrae and embedded tooth are the result of an attack on a live hadrosaur, not the scavenging of a carcass, and represent unequivocal evidence of a predator–prey relationship ( 3, 33). Rugose bone growth encrusting and partially covering the embedded tyrannosaur tooth in the hadrosaur vertebrae indicates that the hadrosaur was unquestionably injured while it was alive and survived the attack long enough to partially heal. The duration of time between the attack and time of death is unknown, however healing rates from modern ectotherms and endotherms suggest that the hadrosaur could have lived from several months to perhaps a number of years after the attack ( 50 – 53). Prey animals that survive attacks by some modern predators, such as lion, hyena, and coyote attacks, frequently can survive long enough to heal ( 27, 55 – 58). Whereas the healed vertebrae demonstrate in this particular case that the attack on the hadrosaur was not consummated by a kill, they are not a sign that T. rex was a poorly skilled predator. Modern terrestrial predators (e.g., coyotes and lions) fail or abort between 45% and 62% of their attacks ( 47, 55 – 58).
Evidence that a T. rex attacked a living hadrosaur indicates predatory behavior, at least in this one instance, and elucidates our view of Western North American Upper Cretaceous paleoecology. Previous assertions that T. rex was exclusively a scavenger would require us to modify our reconstructions of the paleoecology to accommodate a scavenger of such significant proportions. The fact that T. rex engaged in predatory behavior enables a more plausible paleoecological reconstruction, suggesting an ecology similar to those observed today, such as the African savannah ( 27, 58).
Conclusion
We now have conclusive evidence that T. rex indeed engaged in predatory behavior. Whereas previously cited fossil evidence, such as isolated tooth marks, might have been easily misconstrued as the result of scavenging behavior, or predatory behavior carried out by another large theropod, our specimen includes the identities of the prey animal and the attacker and the fact that the prey was alive when attacked. Moreover, the position of the injury—the tail—suggests that T. rex could possibly have engaged in pursuit predation. As a significant component of the Late Maastrichtian Western North American ecology, perspectives of T. rex ’s role as either a scavenger or predator has a profound effect on our view of the paleoecology because it is such a massive animal. As such, our view of this large theropod as a predator enables us to speculate with more confidence on more accurate paleoecological reconstructions for Laramidia in the Late Cretaceous.
Materials and Methods
The vertebrae and tooth crown specimen are reposited at the Palm Beach Museum of Natural History ( PBMNH.P.09.039 ).The fossils were stabilized with cyanoacrylate adhesive PaleoBond PB002 and PB100 and prepared using a Comco MB1000 micro air-abrasive unit with sodium bicarbonate powder. Particularly recalcitrant patches of matrix were removed using a PaleoTools MJ5 micro air-scribe. After preparation, the specimen was sealed with polyvinyl acetate stabilizer and later imaged by CT radiography (Siemens SOMATON Sensation 64-slice). CT scanning revealed the tooth’ s orientation within the vertebrae. The tooth crown is well preserved with enamel and denticles intact.
The morphology and morphometric relationships of the embedded theropod tooth in the present study were examined following the methodology of the Smith et al. ( 46) study. The measurements were compared with the Smith et al. ( 46) data, with large theropod taxa documented in the Hell Creek Formation ( T. rex and Nanotyrannus ), with Albertosaurus as an exemplar of smaller tyrannosaurids.Initially proposed by Bakker et al. ( 47), the taxonomic status of Nanotyrannus has been questioned ( 59) but is considered a valid taxon for this study.
The ratio of two additional sets of measurements for the embedded tooth, DC and ICL, was also examined and compared with the theropod taxa listed above. The DCT is defined in 1cm increments, and the ICL is defined as the specific crown length observed at each of the DCT increments ( Fig. 1 A View Fig. 1 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |