
Oecomys Thomas, 1906
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2001)263<0003:TMOPFG>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03B69D69-FFAB-372E-8503-FB95FCECF9FF |
|
treatment provided by |
Marcus |
|
scientific name |
Oecomys Thomas |
| status |
|
For most of the last three decades, Neotropical mammalogists have identified small specimens of Oecomys (formerly considered a subgenus of Oryzomys ) as O. bicolor and large specimens as O. concolor following Hershkovitz (1960). Unpublished revisionary research, however, suggests that at least 13 valid species of Oecomys are represented among the many nominal taxa that Hershkovitz lumped into bicolor and concolor (see Musser and Carleton, 1993). Four morphologically diagnosable species are known to occur in French Guiana (table 27), of which two are represented among our vouchers. Pending the publication of a comprehensive revision of this difficult genus, we offer preliminary descriptions and diagnostic comparisons of both Paracou species to document our identifications.
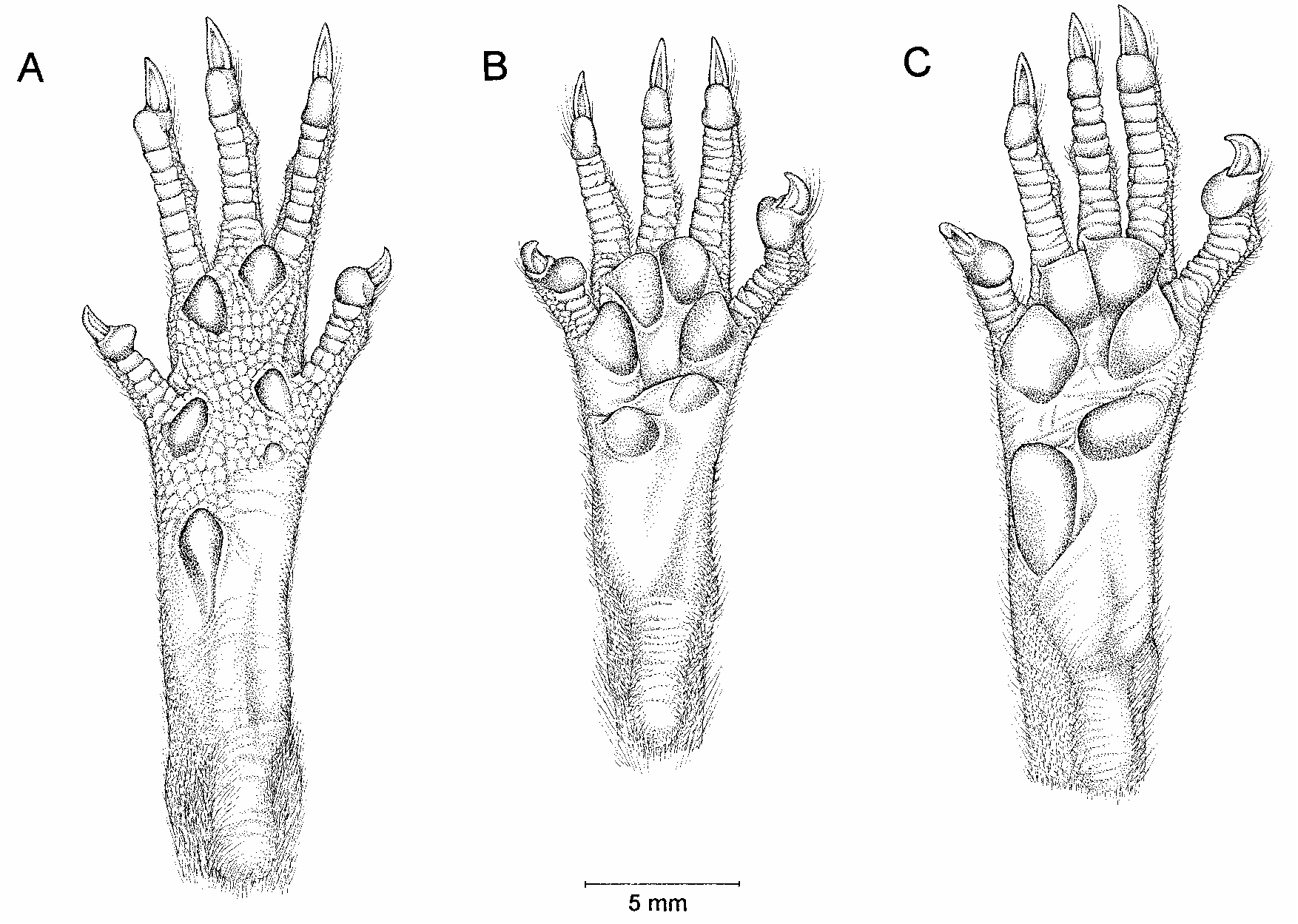
Species of Oecomys are small to mediumsized murids, ranging in average adult body weight from about 20 to 60 g. The dorsal fur is soft (not spiny) and, in adults with fresh pelage, usually some shade of reddish brown. The ventral fur can be either selfcolored (pure white) or graybased with a superficial wash of white, buff, or orange. The mystacial vibrissae are long, extending well behind the pinnae when laid back alongside the head. The dorsal pelage of the hindfeet is sometimes indistinctly darker over the metatarsals than on the digits, but sharply defined metatarsal spots or bands are absent; the plantar surface of the hindfoot is either unpigmented (whitish in preservative, pink in life, brown or amber in dried skins) or lightly pigmented (grayish), but apparently never blackish. Structurally, the hindfeet are short and broad, with large plantar pads and semiopposable fifth digits (fig. 53B). Tails are unicolored in most species (dark above and below), and they are usually longer than the combined length of headandbody; a terminal tuft of long hairs is present in some, but not all species.
Amazonian species of Oryzomys are sometimes misidentified in the field as Oecomys
TABLE 26 Geographic and Morphological Comparisons among Lowland Species of Neusticomys
(and vice versa), but differ externally by their much shorter mystacial vibrissae (not extending behind the pinnae), distinctive hindfeet (fig. 53A), and shorter tails that are often bicolored (at least basally) and never have terminal tufts of long hairs (see the account of Oryzomys below for more detailed descriptions of external traits).
Species of Rhipidomys resemble Oecomys externally in possessing long mystacial vibrissae; shortbroad hindfeet with unpigmented soles, large plantar pads, and semiopposable fifth digits; and long, usually unicolored, tufted tails. However, the hindfeet of Rhipidomys are distinctive, with darker metatarsal markings, larger plantar pads, and relatively longer fifth pedal digits (fig. 53C). In addition, whereas female Oecomys have eight mammae, female Rhipidomys have only six. More detailed comparisons between likesized species of Oecomys and Rhipidomys that might be confused in the field are provided in the account that follows.
Figures 53B View Fig , 55 View Fig , 56 View Fig , 57A View Fig , 62B VOUCHER View Fig MATERIAL: AMNH 266560, 266564, 267593, 267595, 267596; MNHN 1995.1027, 1995.1028. Total = 7 specimens. IDENTIFICATION: Oecomys auyantepui was originally described by Tate (1939) from two specimens collected at 1100 m elevation on
Auyantepui in Estado Bolívar, Venezuela. Hitherto regarded as a junior synonym of O. concolor (by Hershkovitz, 1960), O. trinitatis (by Cabrera, 1961), or O. paricola (by Musser and Carleton, 1993), auyantepui is unambiguously diagnosable from other named forms of Oecomys and merits recognition as a distinct species. Distinguishing traits include its predominantly graybased ventral fur, a distinctly tufted tail, lack of broadly shelved supraorbital margins and postorbital processes, a primitive carotid arterial circulation, presence of an alisphenoid strut, a large postglenoid foramen, complete closure of the subsquamosal fenestra, and a distinctive range of morphometric variation (see table 27 and below). All of the specimens we refer to O. auyantepui are from the Guiana subregion of Amazonia (fig. 54).
In the hand, Oecomys auyantepui is an attractive mouse with soft reddishbrown fur that is much brighter in mature adults with fresh glossy pelage than in juveniles, subadults, or specimens with obviously worn, dull coats. The small ears are covered with a short but macroscopically visible pelage that is colored essentially like that of the head and nape (not contrastingly darker). The ventral fur, superficially whitish, creamcolored, or pale buff, is sharply set off from the reddishbrown fur of the sides and back. The ventral fur is predominantly graybased in most of the specimens at hand, but the fur of the chin and throat is usually selfcolored (all pale), and a few specimens have selfcolored fur extending caudally along the ventral midline to the groin. The hindfeet are either covered uniformly with pale buffy hairs or the metatarsus is indistinctly darker than the toes, but a distinct metatarsal band of blackish fur is apparently never present. Undamaged tails are uniformly dark (almost blackish in some specimens) with a terminal tuft of hairs that are distinctly longer (6–10 mm) than the short (<2 mm) hairs on the proximal part of that organ.
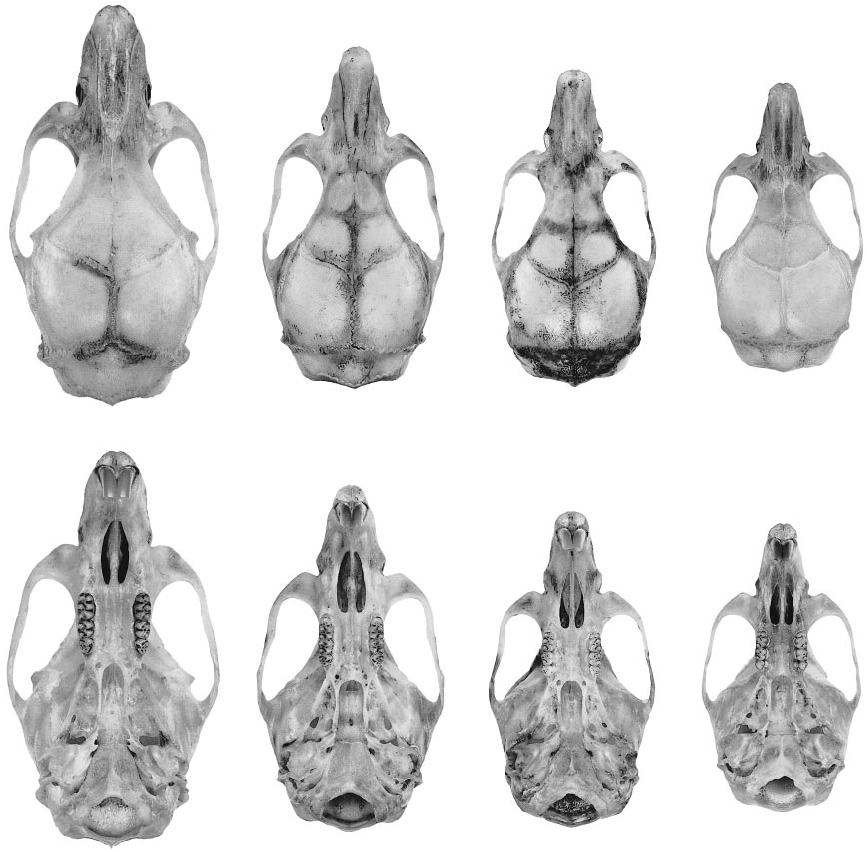
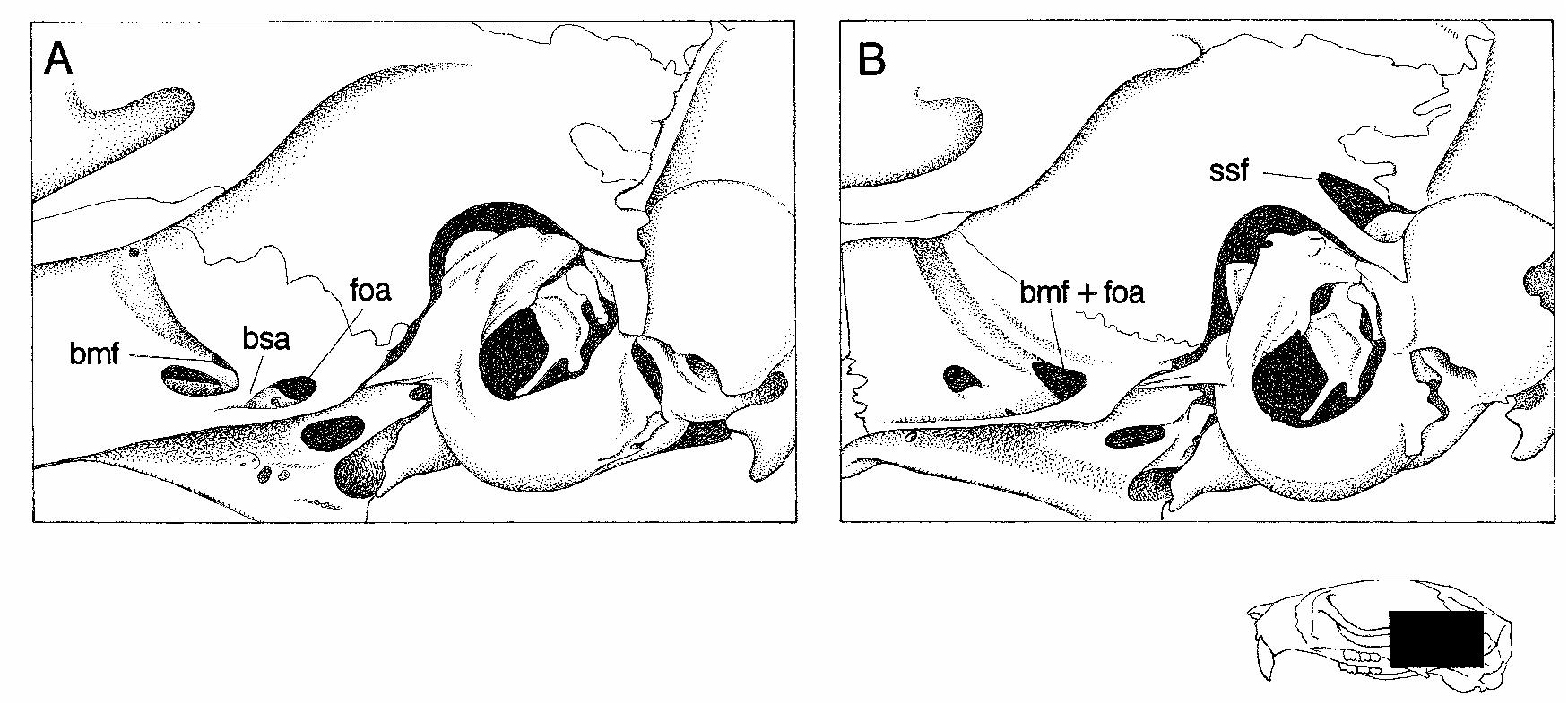
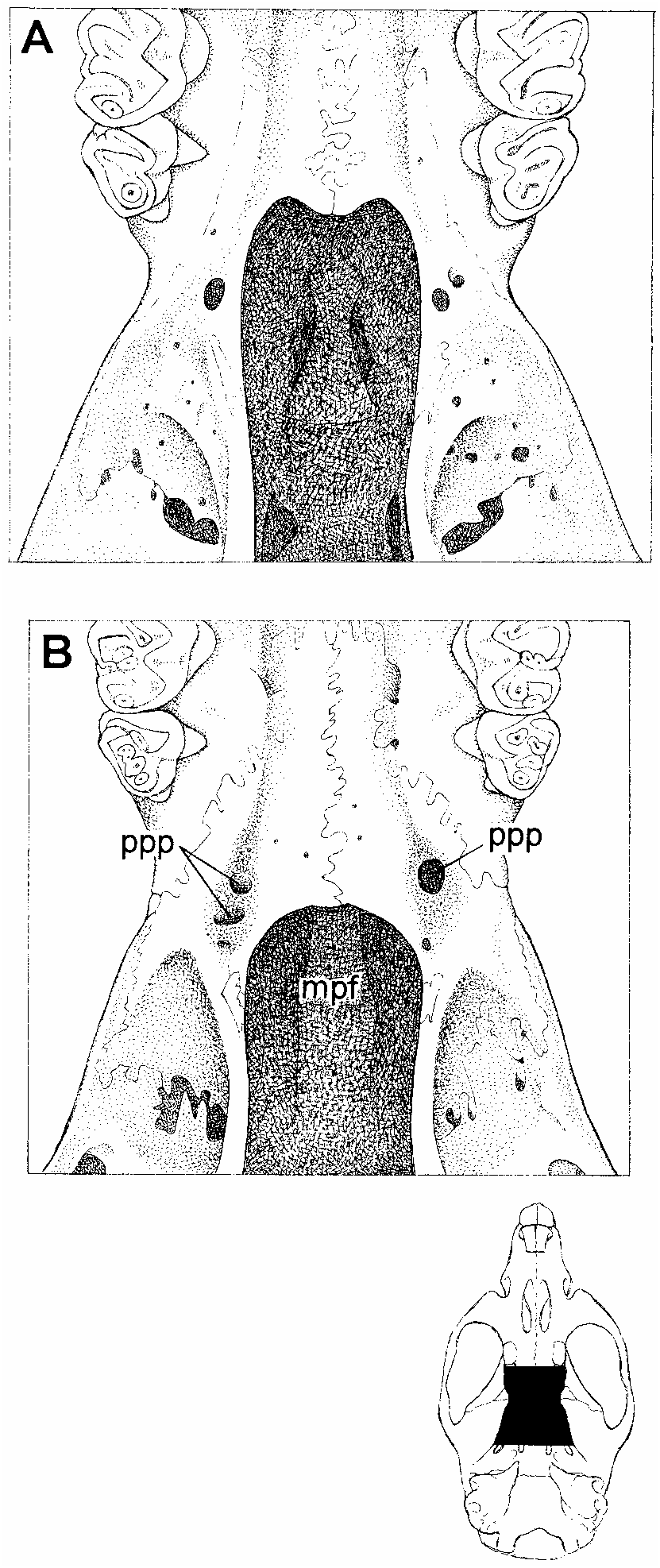
The skull (figs. 55, 56) is unremarkable in general aspect, with the short rostrum, shallow zygomatic notches, convergent interorbit, beaded supraorbital margins, widelong palate, and small bullae characteristic of this oryzomyine genus. The incisive foramina are of average length relative to the diastema (neither very short nor very long by oryzomyine standards), with wide and moreorless evenly convex lateral margins. The roof of the mesopterygoid fossa is completely bony, with no trace of sphenopalatine perforations in most of the specimens at hand. A large stapedial foramen on the medial surface of the bulla, a translucent squamosalalisphenoid groove on the internal surface of the braincase, and a distinct sphenofrontal foramen in the rear of the orbit indicate that the pattern of carotid arterial supply is primitive (pattern 1 of Voss, 1988). A robust diagonal strut of the alisphenoid bone separates the foramen ovale accessorius from the buccinatormasticatory foramen on both sides of the skull in most specimens, and the subsquamosal fenestra is invariably absent (fig. 57A; see table 29).
Morphometrically, series of Oecomys auyantepui that we measured from Guyana, French Guiana, and Brazil (Amapa´) are remarkably similar (table 28). In fact, the most divergent specimen we examined is the Venezuelan type (AMNH 131156), which is larger than most of the other material at hand but does not differ in any qualitative external or craniodental character. Additional Venezuelan material would be useful to determine whether the type represents a western population characterized by large size, or is merely an unusually large individual.
Oecomys auyantepui is intermediate in size to other congeneric species that occur in the Guiana subregion of Amazonia, two of which ( O. bicolor and O. rutilus ) are substantially smaller, and four of which ( O. concolor , O. rex , O. roberti , O. trinitatis ) are larger. Additionally, other Guianan subregion species differ from auyantepui by having entirely selfcolored (pure white) ventral fur ( bicolor , rutilus ), untufted tails ( concolor , rex , roberti , trinitatis ), moreorless bicolored tails ( trinitatis ), broadly shelved supraorbital margins ( rex ), a derived pattern of carotid arterial supply ( concolor ), confluent accessory oval and buccinatormasticatory foramina (most bicolor specimens), and/or patent subsquamosal fenestrae ( bicolor , concolor , rutilus , trinitatis ). Instead, Oecomys auyantepui is morphologically most similar
TABLE 27 Diagnostic Morphological Comparisons among French Guianan Species of Oecomys (Tabulated data include information from specimens collected throughout the Guiana subregion of Amazonia.)
to O. paricola , a species that we do not recognize as occurring in the Guiana subregion.
Oecomys paricola (Thomas, 1904) was originally described on the basis of a very young female specimen (BMNH 4.7.4.63) collected at ‘‘IgarapéAssu’’ (= Igarapé Açu at 1°07̍S, 47°37̍W; Paynter and Traylor, 1991) near Belém. To evaluate the hypothesis that O. paricola and O. auyantepui are synonyms (Musser and Carleton, 1993), we examined the type of paricola and 21 additional specimens collected in the same interfluvial region (south of the Amazon and east of the Rio Tocantins). 15 Because it was not our intention to evaluate the taxonomic status
15 We examined specimens of Oecomys paricola from the following localities: Brazil — Para´, Belem (USNM 393819, 393842, 461386, 545232, 545233), Capim (AMNH 188963, 203391, 203392, 203397; USNM 461385, 461388–461395, 461400), Igarapé Açu (BMNH 4.7.4.63, 4.7.4.64, 4.7.4 108).
of all museum material currently identified as O. paricola , we did not include specimens so determined from other interfluvial regions, some of which exhibit characters not shown by the type.
Oecomys paricola is about the same size as O. auyantepui , and these two species are perhaps indistinguishable in pelage color and external morphology. Cranially, however, two qualitative characters (table 29, fig. 57) permit unambiguous identification. (1) A welldeveloped alisphenoid strut almost always separates the buccinatormasticatory and accessory oval foramina in auyantepui , but an alisphenoid strut is missing and these foramina are consistently confluent in paricola . (2) Whereas the subsquamosal fenestra is consistently absent in auyantepui , a distinct subsquamosal fenestra (separated from the postglenoid foramen by the hamular process of the squamosal bone) is always pre sent in paricola . Although their close resemblance in other respects suggest that auyantepui and paricola are likely to be sister taxa, we recognize them as valid (diagnosable) species allopatrically distributed north and south of the Amazon.
OTHER SPECIMENS EXAMINED: Brazil — Amapa´, Serra do Navio ( USNM 393820 About USNM , 393821 About USNM , 394239–394243 About USNM , 394246–394249 About USNM , 461521 About USNM ) ; Amazonas , 80 km N Manaus ( USNM 579996–580001 About USNM ). French Guiana —Arataye ( MNHN 1986.865 ; USNM 578015 About USNM , 578019 About USNM ), Iracoubo (1983.394, 1983.395), St.Eugène (1994.124, 1995.3235, 1998.1844), Trois Sauts ( MNHN 1983.398 ) . Guyana — Cuyuni Mazaruni, Kartabo ( AMNH 64135 ) ; Potaro Siparuni , 5 km SE Surama ( ROM 102944 About ROM , 103051 About ROM , 103052 About ROM , 103244 About ROM , 103288 About ROM ) ; Upper Demerara Berbice , 18 km SW Kwakwani ( AMNH 269829 , 269830 ), Tropenbos ( ROM 103433 About ROM , 103502 About ROM ) . Venezuela — Bolívar, Auyantepui ( AMNH 131108 , 131156 [holotype]) .
FIELD OBSERVATIONS: All of our definite records of Oecomys auyantepui from Paracou are based on collected specimens. Of
these, one was taken on the ground in a Sherman trap, three were taken in Victor snap traps tied to lianas 1.2–1.5 m above the ground (fig. 58), and three were taken in arboreal platform traps 7.2–10.5 m above the ground. Three specimens were trapped in creekside primary forest, two in welldrained primary forest, and two in swampy primary forest. All specimens were found in the traps at or soon after dawn.
Oecomys rutilus Anthony Figures 55 View Fig , 56 View Fig
VOUCHERS: AMNH 266561, 267584, 267586, 267588–267591, 269121; MNHN
1995.1023–1995.1026. Total = 12 specimens.
IDENTIFICATION: Oecomys rutilus was originally described by Anthony (1921b) based on a single specimen collected by W. Beebe at Kartabo, CuyuniMazaruni District, Guyana. Anthony (p. 4) characterized rutilus as ‘‘A small, brightly colored species, with very short tail and clear white under parts’’, and he remarked that it was quite distinct from another species originally described from Guyana, Oecomys nitedulus Thomas (1910) :
Compared with Oecomys nitedulus , collected at the same place, rutilus is somewhat smaller superficially,
much brighter in color, with longer, softer, pelage, shorter tail and conspicuously smaller skull.
Despite Anthony’s explicit statement that rutilus and nitedulus were valid sympatric species, Hershkovitz (1960) listed both names as synonyms of Oecomys bicolor , a taxon originally described (Tomes, 1860) from eastern Ecuador. Hershkovitz (p. 539) admitted that he had not seen Anthony’s type of rutilus , but remarked that ‘‘Judged by the original description, it is a subadult of the same Kartabo series identified by Anthony as nitedulus . ’’
Oecomys rutilus was listed as a valid species by Musser and Carleton ( 1993), but no account of its diagnostic characteristics has yet been published to supplement Anthony’s preliminary observations. To identify our Paracou material, we examined every available specimen of small Oecomys from the Guiana subregion of Amazonia, including the types of rutilus and nitedulus . In the fol lowing account we provisionally accept the hypothesis that nitedulus and bicolor are conspecific (Hershkovitz, 1960; Musser and Carleton, 1993), but we note that this synonymy remains untested by published analyses of character data, and that we have not made a careful study of typical bicolor (from eastern Ecuador). Instead, our taxonomic comparisons are based exclusively on Guian an material. 16
The frequency distribution of the length of the upper molar row (LM) for small Oecomys collected in the Guiana subregion of Amazonia (N = 66) is distinctly bimodal (fig. 59), with one peak in the interval 3.2– 3.4 mm (including the type of rutilus with LM = 3.26 mm) and another in the interval 3.6–3.9 mm (including the type of nitedulus with LM = 3.79 mm). Although this univariate graph does not of itself show any morphometric discontinuity that could be used to sort specimens into discrete size classes (all frequency intervals between 3.0 and 4.0 mm are occupied), the bimodal pattern clearly indicates that our sample is heterogeneous. Fortunately, variation in other characters is correlated with molar toothrow length and provides unambiguous evidence for species diagnosis. In fact, the specimens we measured of the smaller species, O. rutilus , have molar toothrows shorter than 3.5 mm, whereas our measured specimens of O. bicolor have toothrows longer than 3.5 mm. With larger samples of both species, however, it seems inevitable that the observed ranges of variation in this dimension will eventually be found to overlap.
Oecomys rutilus and Guianan specimens of O. bicolor (= nitedulus ) are similar in external appearance: both are small, usually reddish mice with dark, tufted tails and pure white venters (table 27). Although rutilus averages smaller than bicolor in all standard
16 We examined specimens of Oecomys bicolor from the following localities: Brazil — Amapa´, Serra do Navio ( USNM 393822–393830 About USNM , 393833–393839 About USNM , 393843 About USNM , 394251–394253 About USNM , 461522 About USNM ). French Guiana —Les Nouragues ( AMNH 269823 ; V882, —895, —896, —898, —912), Rivière Approuague ( MNHN 1983.393 ). Guyana — Cuyuni Mazaruni, Kartabo ( AMNH 41910 , 60641 , 64130 , 64132 , 64133 ) ; Potaro Siparuni , 5 km SE Surama ( ROM 102970 About ROM ) ; ‘‘ Essequibo River 13 mi from mouth’’ ( BMNH 6.4 .8.31 [holotype of nitedulus ]; ‘‘ R. Supinaam’ ’ ( BMNH 10.9.29.18).
TABLE 28 Measurements (mm) and Weights (g) of Adult Oecomys auyantepuia TABLE 29 Comparisons of State Frequencies for Two Qualitative Craniodental Characters in Oecomys auyantepui and O. paricola a
external dimensions (table 30), there is sufficient morphometric overlap between them that no measurement is diagnostic. Most of the material at hand is preserved in fluid, so it is difficult to evaluate Anthony’s statement that these species differ in fur color; however, no consistent differences were apparent in the few dried skins we examined. The species difference in tail length mentioned by Anthony (presumably meant to be considered in relation to the headandbody) is also difficult to assess because we have not measured any specimens of bicolor ourselves (minor differences in measurement methodology can produce substantial artifactual divergence in computed ratios). Instead, two other characters are useful for field identification.
As observed by Anthony, the dorsal fur is longer in Oecomys rutilus than it is in O. bicolor , and the two species also differ in fur texture. In rutilus , the fur averages about 6– 8 mm middorsally near the rump, and it feels soft and lax when ruffled because the unresistant hairs return slowly to their normal (unruffled) condition. By contrast, the dorsal fur of bicolor is only 4–5 mm long in most specimens, and because bicolor is the larger species this absolute difference makes the pelage appear relatively much shorter. Also, the short fur of bicolor feels ‘‘crisp’’ to the touch because the stiffer hairs are more resistant to ruffling.
The tuft of hairs at the tailtip is significantly longer in Oecomys rutilus , averaging 7.7 ± 1.4 mm (observed range: 5–11 mm, N = 19). In O. bicolor , this tuft measured 4–5 mm in all nine specimens we examined with intact tail tips. Since the long hairs of the tail tuft are often exposed to bending and compression in museum trays, it is probable that some of the tufts we measured were broken short, so the mean tuft length determined from fresh specimens with undamaged tails might be larger for both species than the values reported here. Nonethless, the difference is visually conspicuous and is useful for sorting skins in combination with other traits.
Skulls of Oecomys bicolor and O. rutilus differ in size (figs. 55, 56; table 30) and in the usual sizecorrelated proportions, but the incisive foramina are notably longer relative to the diastema in bicolor than in rutilus , a difference that is not attributable to standard patterns of muroid craniodental allometry. Although bicolor and rutilus are similar in most qualitative osteological traits, they differ significantly in the frequency of occurrence of the alisphenoid strut, an ossification that occurs bilaterally in most rutilus , but is bilaterally absent in most bicolor (table 31). Otherwise, these species are craniodentally similar, both having primitive carotid circulations (pattern 1 of Voss, 1988), beaded but unshelved supraorbital margins, and consistently large postglenoid foramina and subsquamosal fenestrae.
Although Oecomys rutilus and O. bicolor are widely distributed in the Guiana subregion of Amazonia, they have been collected
TABLE 30 Measurements (mm) and Weights (g) of Adult Oecomys rutilus and O. bicolor a
sympatrically only at Kartabo (CuyuniMazaruni District, Guyana) and Les Nouragues (French Guiana). Musser and Carleton ( 1993) gave the range of rutilus as restricted to Guyana, Surinam, and French Guiana, but material from San Ignacio Yuruaní ( Venezuela) and from 80 km N Manaus ( Brazil) previously misidentified as bicolor (e.g., by Voss, 1991: table 23; Voss and Emmons, 1996: appendix 7) extend the range of this species east and south of the Guianas proper (fig. 60). Although we have not examined any material of rutilus from outside the Guiana subregion of Amazonia, the small unnamed Oecomys that Patton et al. (2000) reported from the Rio Juruá appears to be similar in some respects and merits close comparison in any future revisionary study.
The material we examined of Oecomys rutilus is remarkably uniform with little indication of significant variation among samples collected at widely separated localities. Thus, measurements of the Guyanese type (AMNH 42910), a mature adult female (not a subadult as conjectured by Hershkovitz, 1960), are all within the range of variation exhibited by French Guianan specimens (table 30). A few measurements of Brazilian and Venezuelan specimens exceed the observed range of variation for homologous measurements of French Guianan material, but the discrepancies are small in all cases.
OTHER SPECIMENS EXAMINED: Brazil — Amazonas, 80 km N Manaus ( USNM 579992–579995 About USNM ). French Guiana —Cacao ( MNHN 1983.400 ), Les Nouragues ( AMNH 269822 , V889, —892, —899, —900, —906, —913), St. Eugène ( MNHN 1995.3236 , 1995.3237 , 1998.1845 , 1998.1846 ) . Guyana — Cuyuni Mazaruni, Kartabo ( AMNH 42910 [holotype], 142820) ; Upper Demerara Berbice, Dubulay Ranch ( AMNH 267745 ), 18 mi SW Kwakwani ( AMNH 269828 ), Tropenbos ( ROM 103482 About ROM ) . Surinam — Suriname, Carolinakreek ( FMNH 95591 About FMNH ) . Venezuela — Bolívar, San Ignacio Yuruaní ( AMNH 257268 , USNM 448576 About USNM ) .
FIELD OBSERVATIONS: Although we saw small reddish mice racing along branches or lianas at night on many occasions, all of our definite records of Oecomys rutilus at Paracou are based on collected specimens. Of our 12 vouchers, 7 (58%) were taken in pitfall traps, 2 (17%) were shot at heights of 4–5 m in trees, 1 (8%) was taken in a Victor snaptrap tied to a liana 2 m above the ground, 1 was taken in a Sherman trap placed on a liana 30 cm above the ground, and 1 was taken in a platform trap 15.9 m above the ground. Six specimens (50%) were taken in welldrained primary forest, 4 (33%) in swampy primary forest, 1 (8%) in creekside primary forest, and 1 in roadside secondary growth. Both shot specimens were taken at night, and all of the other specimens were found in the traps at dawn.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |