
Macrocentrus pryeri Yang, Song & Cao, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4303.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:2BE0876D-5512-460A-85D1-213D913A1CBC |
|
DOI |
https://doi.org/10.5281/zenodo.6008989 |
|
persistent identifier |
https://treatment.plazi.org/id/03B18671-FFCC-481B-22C7-FE6216B48C89 |
|
treatment provided by |
Plazi |
|
scientific name |
Macrocentrus pryeri Yang, Song & Cao |
| status |
sp. nov. |
Macrocentrus pryeri Yang, Song & Cao , sp. nov.
( Figs 5–20 View FIGURES 5 – 8 View FIGURES 9 – 12 View FIGURES 13 – 16 View FIGURES 17 – 21 )
Type material. Holotype, ♀, Seed orchard of Pinus koraiensis, Lushuihe Forest Bureau , Jilin Province, June 20, 2016, reared from larvae of Dioryctria pryeri attacking twig of Pinus koraiensis by Li-wen Song, Xing-peng Li and Zhong-qi Yang. Paratypes, 60♀♀ 26♂♂, Topotypic, and same data as holotype.
Description. Female.
Body length. Holotype body length 6.3 mm, fore wing 5.3 mm; antenna 1.95 times as long as fore wing.
Head ( Figs 5–7 View FIGURES 5 – 8 , 9 View FIGURES 9 – 12 ). Antennae with 48 antennomere ( Fig. 10 View FIGURES 9 – 12 ), long setae, length of third antennomere 1.28 times fourth antennomere, Length of third, fourth and penultimate antennomere 8.0, 6.3, 2.9 times their width respectively; apical antennomere with a distinct spine; length of maxillary palp 1.4 times height of head; maxillary and labial palpi with 6 and 4 segments, respectively; length of third segment of labial palp distinctly shorter than fourth segment and only 0.55 times the latter; length of eye in dorsal view 2.8 times temple ( Fig.7 View FIGURES 5 – 8 ); temple gradually narrowed posteriorly; OOL:OD:POL=14:9:10; frons flat, smooth and shining without crenulae; vertex moderately convex and outside of ocellar area smooth with dense setae and not punctulate laterally; face convex medially, smooth and with dense, even setiferous punctations; clypeus moderately convex medially with ventral margin slightly concave, and with sparser setae than on face ( Fig. 9 View FIGURES 9 – 12 ); length of malar space 0.67 times basal width of mandible; mandible twisted apically, its second tooth robust, distinctly shorter than first one, and rounded apically; upper level of mandibular condyli below level of eyes.
Mesosoma ( Figs 5–8 View FIGURES 5 – 8 , 11 View FIGURES 9 – 12 ). Length of mesosoma 1.45 times its height; no distinct pronope; side of pronotum coarsely rugose medially and postero-ventrally, finely rugose medio-anteriorly, remainder largely smooth, pronotum with anterior margin sharp-carinated and with relatively dense setae medio-dorsally; mesopleuron convex, with relative dense setae and small setiferous punctations; subalar tubercle convex; prepectal carina very weak and only developed ventrally with shallow groove behind carina ( Fig. 11 View FIGURES 9 – 12 ), its lower part extending backwards for 1/3 length of the carina and; anterior margin of prepectus with a crenulate sulcus which extends upwards and contacted to shallow trough-like epicnemial depression, and its anterior-upper corner rugulose; mesopleural fovae shallow; precoxal sulcus not distinctly developed; mesopleural suture extending upwards to anterior subalar depression, both sulci striate-rugose; mesosternal sulcus shortly developed posteriorly; postpectal carina sharp; metapleuron moderately rugulose with dense setae which are distinctly longer than those on mesopleuron; metapleural flange medium-sized, obtuse, apically upcurved; mesoscutal lobes smooth and shining; middle lobe convex, with dense setae; lateral lobes slightly convex, with sparse setae, but median area bare; scutellum moderately convex, with prominent median-posterior margin, its anterior-median area having sparse and short setae and remainder bare, smooth, scutellar sulcus large, two pits, finely rugose inside ( Fig. 7 View FIGURES 5 – 8 ); metanotum with distinct median carina and two sublateral carinae, but median carina only at anterior 3/5 and median-posterior part rhomboid, smooth; the median area between sublateral carinae superficially rugose; lateral parts of metanotum coarsely rugose; propodeum convex, without median carina, and densely rugose, with setae longer and denser laterally, sparser and shorter dorsally, subposterior-median area bare ( Fig. 7 View FIGURES 5 – 8 ).
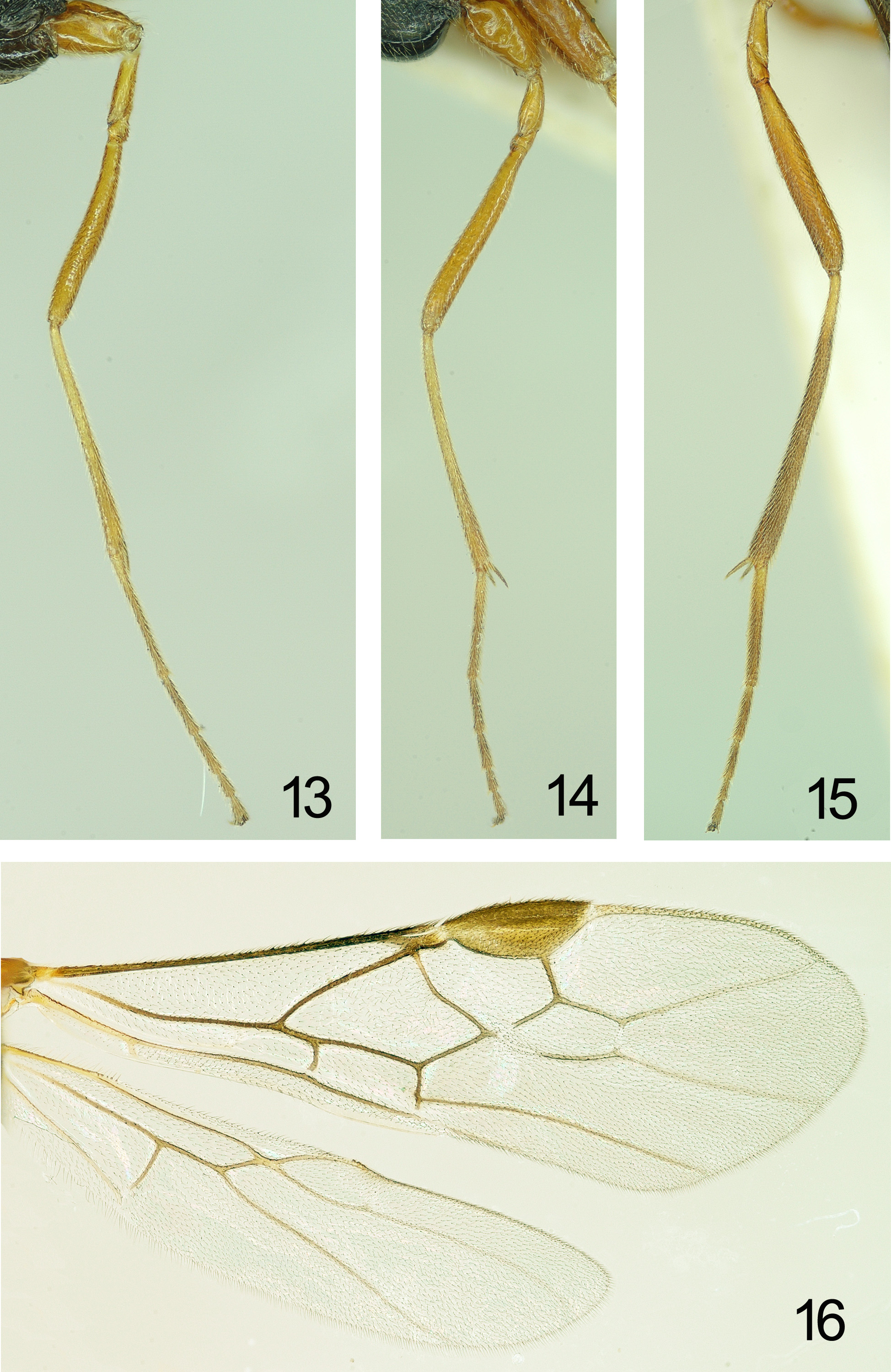
Wings ( Figs 5, 6 View FIGURES 5 – 8 , 16 View FIGURES 13 – 16 ). Fore wing: for marginal cell, see Fig. 16 View FIGURES 13 – 16 ; subbasal cell sparsely setose basally and lower 1/3 bare, lacking pigmented patch; r:3-SR:SR1=12:18:58; 1-CU1:2-CU1=11:30 (2.72 times); 1-CU1 widened, about 1.5 times as broad as 2-CU1, and nearly as long as CU-a; CU-a slightly curved outwards, subparallel with 3- CU1 ( Fig. 16 View FIGURES 13 – 16 ); 2-SR:3SR:r-m=22:20:8; 2A largely sclerotized, membraneous base sparsely setose. Hind wing: 3 hamuli; SC+R1 slightly curved; marginal cell parallel-sided apically; 1-M 1.5 times 1r-m ( Fig. 16 View FIGURES 13 – 16 ).
Legs ( Figs 13–15 View FIGURES 13 – 16 ). Hind coax nearly completely smooth, with some micro-sculpture posteriorly and without striae; tarsal claw slender without basal lobe and bristly setose; fore femur slender and evenly curved, but slightly wider medially than base and apex; length of fore spur 0.26 times basitarsus; length of femur, tibia and basitarsus of hind leg 8.0, 11.8 and 9.5 times their width, respectively; with two spurs, in which one is 0.37 times and another is 0.42 times hind basitarsus length, respectively; left fore, middle and hind trochantelli with 7, 6 and 6 small teeth respectively, however, right fore trochantellus with 8 teeth.
Metasoma ( Figs 5–8 View FIGURES 5 – 8 , 12 View FIGURES 9 – 12 ). Length of first tergite 1.9 times its apical width ( Fig. 12 View FIGURES 9 – 12 ), its surface finely and densely longitudinally striate, medio-basally distinctly concave, dorsal carinae of first tergite absent; second and third tergites finely longitudinally striate, but third tergite with apical 1/10 and apical lateral areas smooth; fourth tergite finely longitudinally striate on basal 1/3; remainder of metasoma smooth; all tergites relatively densely setose; length of ovipositor sheath 1.54 times fore wing and 1.23 times body length; apex of ovipositor rather slender, with small notch subapically.
Color. Body entirely black; clypeus, scapus, pedicellus, basal half of third segment of antenna, palpi, tegulae, all legs slightly brownish yellow, but apices of tibiae and tarsi slightly infuscate; remainder of antenna, apex of scutellum, ovipositor sheath dark brown; veins and pterostigma light brown, but veins at apical area after pterostigma of fore wing and veins of hind wing pale yellow; wing membrane subhyaline.
Variation. Number of left and right antennal segments of female in the same individual sometimes differ, from 48 to 49 respectively; third segment of antenna sometimes with color identical to following antennal segments; fourth tergite with striae at basal 1/3 area sometimes very superficial, but distinct.
Male ( Figs 17–21 View FIGURES 17 – 21 ). Generally similar to female, but body length shorter, 5.0– 5.8 mm; fore wing length 4.5– 5.0 mm; antenna with all flagellum segments dark brown; fore wing with pterostigma light-brown yellow; left antennal segments variable, 44–46, right antennal segments variable, 43–46; third tergite with longitudinal striae only at basal 1/3 and remainder area smooth; basal area fourth tergite without superfacial striae.
Distribution. China ( Jilin Province).
Etymology. The specific epithet is derived from its host species name “ pryeri ” ( Dioryctria pryeri ).
Diagnoses. The new species is close to M. sylvestrellae Achterberg (2001) which is a parasitoid of Dioryctria sylvestrella , but differs from the new species as follows: tergite 1–3 with distinct longitudinal striations, tergite 4 finely but obviously longitudinally striate on basal 1/3 ( Fig. 12 View FIGURES 9 – 12 ); frons with some crenulae; outside of ocellar area punctulate laterally; face unevenly convex, coarsely punctate, rather densely so medio-ventrally and sparsely so laterally, and medio-dorsally with weak rugae; side of scutellum rugose; fore wing with subbasal cell having faintly indicated pigmental patch, vein 2-CU1 4.6 times 1-CU1 and 1-CU1 not widened; hind wing with vein 1-M 1.9 times 1r-m. The new species is also very close to another Chinese Macrocentrus species, M. watanabei Achterberg (Achterburg 1993b) , a parasitoid of Dioryctria rubella , but can be separated from the latter by tergite 4 which is finely but obviously longitudinally striate.
Biological Observations. Macrocentrus pryeri sp. nov. is a gregarious endo-parasitoid of larva of Dioryctria pryeri Ragonot ( Lepidoptera : Pyralidae ) on Pinus koraiensis . Probably it is a polyembryonic as other gregarious parasitoid species of Macrocentrus ( Shaw and Huddleston 1991) . In Changbaishan forest areas, Jinlin Province, its host D. pryeri had a single generation per year but with a span over two years for developing and finishing the generation ( Mao et al. 2006, Song 2016, Zang 2007, Zhang and Ge 2011). The moth emerged from the second ten days of June to the first ten days of July and the female laid eggs on second year green cones. After hatching the larvae entered cones, mining inside and seriously damaging the seeds and cone tissues. Usually in the 2nd ten days of September the 4th or 5th instar larva emerged from the cones and moved to the base of 2nd year old lateral shoots, entering the shoot by mining in the phloem, the tree releasing resin to form a resin-gall. The mature larva later overwintered under the resin-gall. At the beginning of May of the following year, the overwintered larvae started to tunnel in the shoots with more resin released outside the tunnel until late June ( Figs 1, 2 View FIGURES 1 – 4 ). The larvae then pupated in the tunnel end under the resin-gall.
Macrocentrus pryeri developed two generation per year ( Song 2016). It parasitized its host at 3rd instar larval stage (in cone-attacking period), and mature larval stage (in shoot-attacking period). We collected the pyralid larvae in the shoots in the late May and the early June for rearing. Mature parasitoid larvae were found surrounding the mummy of their host larva. They then spun slightly yellow-brown silk cocoons which were tightly connected similar to honeybee comb in cross section ( Figs 3, 4 View FIGURES 1 – 4 ).
The natural parasitism rate of the gregarious parasitoid ranged from 15–40%, with an average of 27.5% for the mature larval stage. From a single host, 29–48 wasp individuals with an average of 40.4 were produced and a female:male ratio of 1.9:1 (maximum 2.8:1). Macrocentrus pryeri appears to have potential as a biological control agent for suppression of its hosts because of its relatively high parasitism rate, the relatively large number of wasp individuals reared from a single host pupa, and the high female:male sex ratio. We have successfully reared the new parasitoid, so mass rearing should be feasible.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |