
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/ 10.1126/science.1138709 |
|
DOI |
https://doi.org/10.5281/zenodo.3809974 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF87C4-FFB4-FF87-FF6A-DBB5F9BDADF1 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
It has long been assumed that the process of fossilization results in the destruction of virtually all original organic components of an organism,and it has been hypothesized that original molecules will be either lost or altered to the point of nonrecognition over relatively short time spans (well under a mil lion years) ( 1 — 7). However, the discovery of intact structures retaining original transparency, flexibility, and other characteristics in speci mens dating at least to the Cretaceous ( 8, 9) suggested that, under certain conditions, rem nant organic constituents may persist across geological time. The skull, vertebrae, both femora and tibiae, and other elements of an exceptionally well- preserved Tyrannosaurus rex [ MOR 1125 ( 5)] were recovered from the base of the Hell Creek Formation in eastern Montana (USA), buried within at least 1000 m3 of medium-grained, loosely consolidated sandstone interfingered with fine-grained muds, interpreted as stream channel sediments. Demineralization of femur and tibia fragments revealed the preservation of fibrous, flexible, and apparently original tissues, as well as apparent cells and blood vessels ( 5), but the endogeneity and composition of these structures could not be ascertained without further analyses. We present molecular and chemical ( 10) analyses of tissues remaining after partial demineralization (11) of the left and right femora and associated medullary bone (72) that would, in extant bone, represent the extracellular matrix (osteoid) dominated by collagen I ( 13). Because of its ordered structure as a triple helix (14, 15), collagen I has unique characteristics that are highly conserved across taxa, making validation of its presence relatively straightforward. The molecular composition of collagen incorporates glycine, the smallest amino acid, at every helical turn. Therefore, an amino acid profile of collagen results in -33% glycine content ( 14). This molecular structure also results in packing of microfibrils with a banded repeat of ~70 nm ( 15, 16). Collagen also shows posttranslational hydroxylation of about half of all proline and some lysine residues; thus, the detection of hydroxyproline and hydroxylysine in extracts of oiganic material is viewed as strong evidence for the presence of collagen (17, 18). Finally, collagen is identified by polyclonal or monoclonal antibody reactivity that can distinguish between collagen types ( 19). We focused on identifying collagen-like compounds because in addition to being abundant and easily identified by multiple and independent methods, this protein is durable ( 20, 21) and resistant to degradation.
The fibrous nature of demineralized dinosaur tissues was demonstrated by optical ( 8) and electron (fig. S1) microscopy. Furthermore, regions of dinosaur cortical and medullary ( 12) bone demonstrated a repeat pattern with periodicity of ~70 nm when examined by atomic force microscopy (AFM) ( Fig. 1, A to D View Fig. 1 ), consistent with collagen in extant bone ( Fig. 1, E and F View Fig. 1 ) and similar to that previously observed in fragments of demineralized Cretaceous avian bone ( 22). However, this periodic pattern was rarely observed in ultrathin sections of MOR 1125 demineralized bone by transmission electron microscopy (TEM) (fig. S1). This may be a methodological problem, or the periodic features we observe ( Fig. 1, A to D View Fig. 1 ) may be due to surface features generated when demineralization removed most of the apatite crystals emplaced during biomineralization, when collagen acted as a template. Thus, the banded features may represent a type of natural molecular imprinting ( 23), because banded fibers have been observed by TEM for other dinosaur tissues ( 24).
TEM studies confirm that, unlike extant bone, dinosaur bone did not completely demineralize after prolonged incubation in EDTA (11). Selected-area electron diffraction (SAED) of the tissues (fig. S1D, inset) showed that this retained mineral is biogenic hydroxylapatite ( 25). It is not possible to determine this conclusively because of the similarity in structure between hydroxylapatite and fluorapatite; however, the observed diffraction circle intensities are most consistent with hydroxylapatite. This finding suggests that the bone mineral is virtually unchanged from the living state and has undergone little if any alteration.
Force curve measurements of demineralized dinosaur medullary and cortical bone indicate that the elasticity of dinosaur tissues was similar to that of demineralized extant bone. We measured both embedded sections (fig. S2A) and unembedded whole mounts (fig. S2B) of demineralized bone in both air and liquid (11). The demineralized bone surface softened after exposure to buffer, allowing the AFM tip to penetrate deeper into the tissues with less resistance. Thus, the modulus of elasticity (fig. S2C) was reduced in liquid by more than three orders of magnitude (fig. S2B). Although ~2000 nN of force was required to penetrate ~40 nm into MOR 1125 bone matrix in air, only ~15 nN of force was required to depress the tip ~75 nm into the same matrix when hydrated (fig. S2B, inset).
MOR 1125 cortical and medullary wholebone extracts showed reactivity to antibodies raised against chicken collagen I (11) when measured by enzyme-linked immunosorbent assay (ELISA), although the degree of binding varied widely. Reactivity was greatly reduced in dinosaur extracts relative to extant samples (fig. S3), but still at least twice that observed in negative controls of coextracted sediments and buffer without sample, similarly treated.
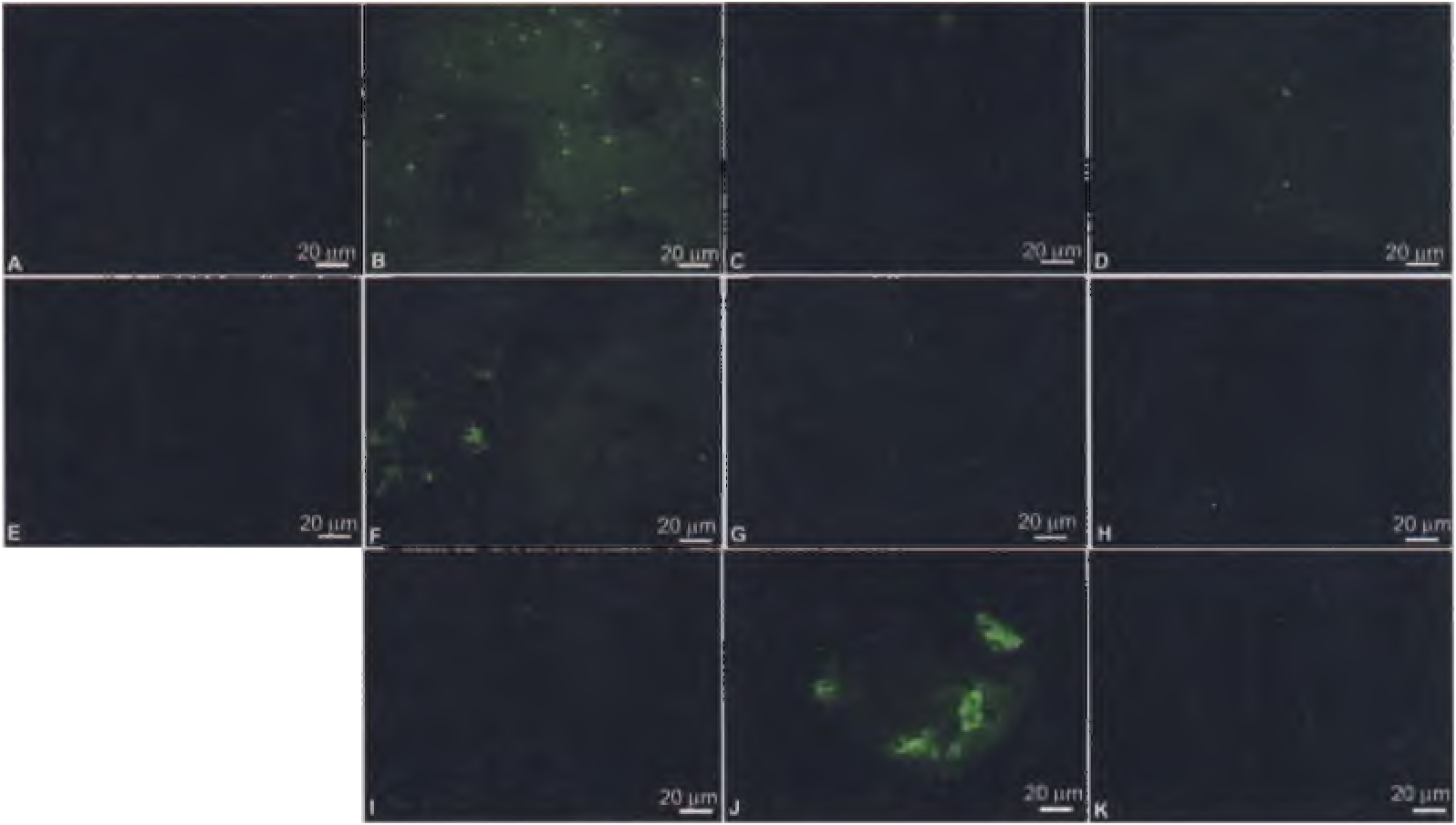
We confirmed the antibody reactivity data by in situ immunohistochemistry in a series of experiments. We exposed thin (0.3 to 0.5 pm) sections of demineralized cortical ( Fig. 2, A to D and I to K View Fig. 2 ) and medullary ( Fig. 2, E to H View Fig. 2 ) dinosaur bone to antibodies raised against avian collagen I, both before ( Fig. 2, B and F View Fig. 2 ) and after inhibition of antibodies with chicken collagen ( Fig. 2, D and H View Fig. 2 ) (11). Additionally, antibody reactivity ( Fig. 2J View Fig. 2 ) was significantly decreased after we digested dinosaur tissues with collagenase ( Fig. 2K View Fig. 2 ), although this enzyme effect was not consistently observed. Reactivity to antibodies, measured by fluorescence, was significantly greater than in negative controls ( Fig. 2, A, C, E, G, and I View Fig. 2 ) and was localized to tissues. We also observed some binding of osteocalcin antibodies to dinosaur tissues (fig. S4). These patterns were similar to those observed with extant emu cortical and medullary bone (fig. S5). Immunoreactivity in dinosaur tissues was greatly reduced from that observed in extant bone, as illustrated by longer integration times and fainter signal, but was greater than in negative controls. Immunohistochemistry performed on sediments was negative for binding. These results imply that the concentration of reactive epitopes in the dinosaur tissues is very low, consistent with the ELISA results. That antibody reactivity was more consistently observed in situ than in ELISA could be due to greater alteration and/or loss of organic compounds during extraction procedures, or to reduced binding of degraded antigen to ELISA plate polymers.
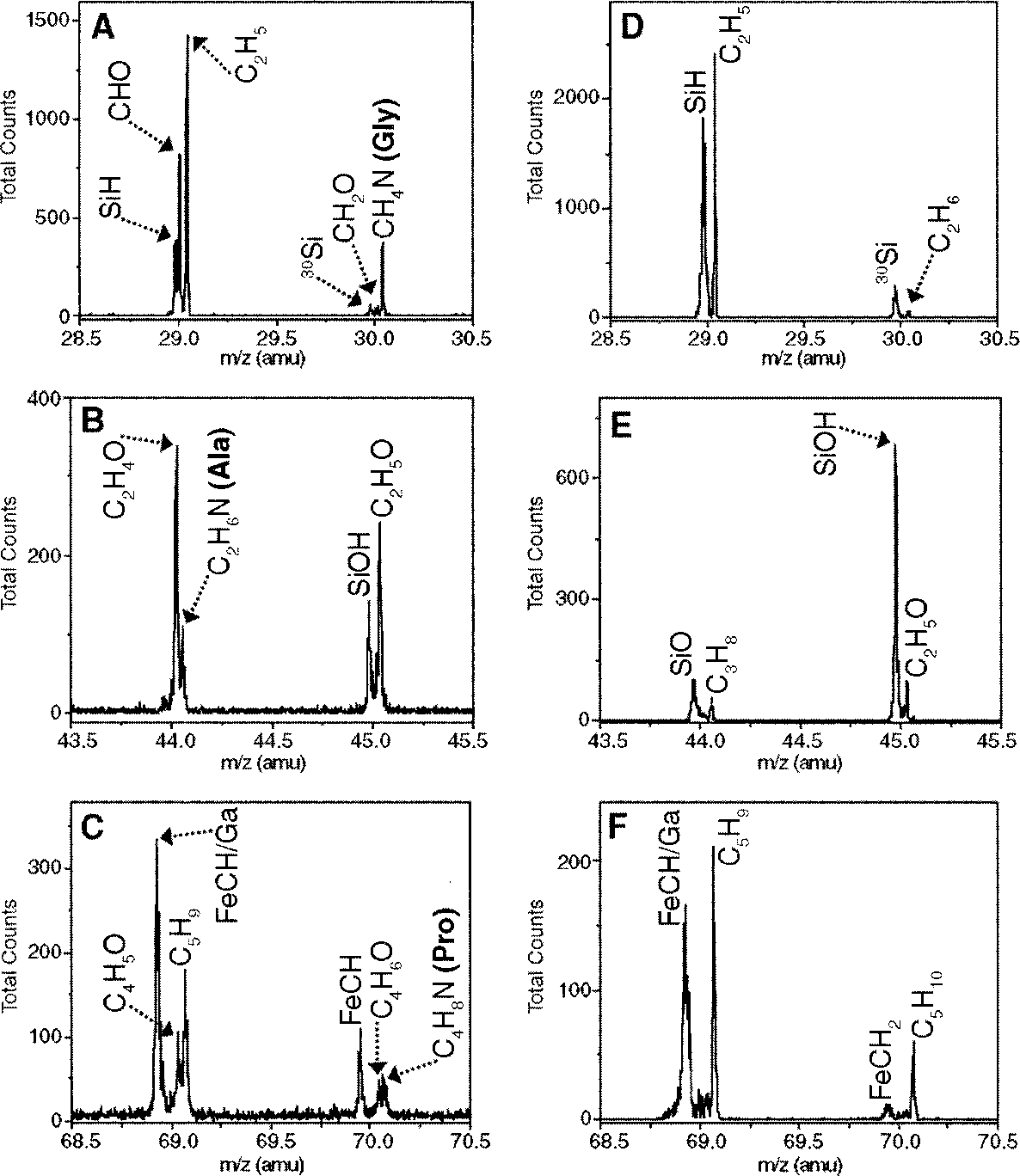
The presence of collagen-derived epitopes in demineralized tissues is supported by mass spectrometry data. Time-of-flight secondary ion mass spectrometry (TOF-SIMS) detects surface ions associated with molecular fragmentation with high mass resolution, and can localize signal to whole samples without subjecting them to chemical extraction. In situ TOF-SIMS analyses were performed to unambiguously detect amino acid residues consistent with the presence ofprotein in demineralized MOR 1125 tissues ( Fig. 2 View Fig. 2 and fig. S6). We obtained ratios of glycine (Gly), the most abundant amino acid in collagen [-33% (74)], and alanine (Ala), which constitutes about 10% of collagenous amino acids, to support the presence of the specific collagen α1 type 1 protein in these tissues. Small peaks representing proline (Pro) at mass/charge ratio (m/ z) 70 ( Fig. 3C View Fig. 3 ), lysine (Lys) at m/ z 84 (fig. S6A), and leucine or isoleucine at m/ z 86 (fig. S6B) were also detected. TOF-SIMS is highly matrix dependent, and desorption and ionization of some amino acid residues, especially modified residues such as hydroxylated Pro, are less efficient than for other residues ( 26). These modified residues were not detected by this method but were readily identified by other mass spectrometry methods ( 10).
The Gly:Ala ratio for published chicken collagen al type 1 sequence (27) is 2.5:1. The TOF-SIMS results show that the Gly:Ala ratio in medullary bone of MOR 1125 is 2.6:1 ( Fig. 3, A and B View Fig. 3 ). Sandstones entombing the dinosaur, subjected to TOF-SIMS as a control, showed little or no evidence for these amino acids ( Fig. 3, D and E View Fig. 3 ).
We identified a variety of nitrogen-containing species—including an alkyl amine group, C7H18N2+, located at 130 amu (fig. S6C)—in all dinosaur samples tested, but not in any surrounding sediments. We also observed a number of Fe-C-H species such as FeCH, FeCH2, and FeCH3, associated with the dinosaur matrix (fig. S7) but not seen in extant material. Similar compounds were observed in the sediments surrounding the dinosaur. These may be microbial products, as sequences from iron-containing microbial enzymes were identified by mass spectrometry ( 10). We interpret these fragments as evidence that iron may help preserve soft tissue through initiation of intra- and intermolecular cross-links ( 9).
Dinosaur protein sequence, including collagen, should be most similar to that of birds among extant taxa, according to other phylogenetic information ( 28). The hypothesis that molecular fragments of original proteins are preserved in the mineralized matrix of bony elements of MOR 1125 is supported by peptide sequences recovered from dinosaur extracts, some of which align uniquely with chicken collagen al type 1 ( 10).
The amount of protein or protein-like components in MOR 1125 is minimal. The percent yield after extraction and lyophilization was ~0.62% for cortical bone and 1.3% for medullary bone. Protein-derived material is only a small percentage of the lyophilate relative to other material coextracted from bone, as assessed by comparison of immunoreactivity with extant samples. This is verified by mass spectrometry, which identifies only femtomole amounts of sequenceable material ( 10) in a heterogeneous mixture of extracted material.
Microenvironments within a single bone vary greatly, and not every fragment ofbone examined yielded positive results. There was a high degree of variability between extractions, and we have also noted progressive reduction of signal in more recent extractions, indicating bone degradation in modem environments ( 29). Therefore, each ofthe analyses we report has been repeated numerous times, and we have set a minimum of three repetitions with similar results before reporting an assay as positive. Additionally, experiments have been conducted independently in at least three different labs and by numerous investigators, and the results strongly support the endogeneity of collagen-like protein molecules.
We hypothesize that these molecular fragments are preserved because reactive sites on the original protein molecules became irreversibly cross-linked, both to similar molecules and to mineral or exogenous organic components. These cross-linking reactions may have been initiated by unstable metal ions that formed free radicals ( 30, 31), which in turn reacted with organic molecules to form polymers (6, 7, 9, 32). We propose that the unstable metal ions were derived from the post mortem degradation of iron-containing dinosaur biomolecules such as hemoglobin, myoglobin, and possibly cytochromes ( 9, 31). Once stabilized by these cross-linking reactions, the molecules were no longer available as substrates for further degradative reactions.
The intimate relationship between apatite and the organic phase of bone also contributes to the preservation oforganic matter (16, 33-38), but we propose that the mineral phase may be stabilized by this relationship as well. The presence of biogenic apatite in these 68-million-year-old bones can only be rationalized by protection from an intact organic phase, which in turn is only satisfied by a synergistic relationship between collagen and mineral phases. Whereas extant bone retains no detectable calcium after days to weeks of demineralization, dinosaur bone retains a fraction of recognizable apatite crystals after months of treatment (fig. S1). Another contributing factor in the retention of original mineral may be that apatite is stabilized in the presence of calcite ( 33).
Sandstones surrounding MOR 1125 contain abundant calcite cements.
The depositional environments may affect organic preservation in other ways. Comparison offossils from a variety of environments indicates that those derived from sandstones are more likely to retain soft tissues and/or cells ( 9). We hypothesize that the porosity of sandstones may facilitate draining of enzymes of decay and suppurating fluids as the organism degrades, whereas organisms buried in nonporous mudstones or clays may be exposed to these longer and therefore may be more completely degraded.
Our findings indicate the need for optimizing methods of extraction and handling of fossil material. In particular, the decrease in signal we observed over time supports the need to establish field collection and storage offossils according to protocols that allow future analytical studies ( 29).
The data presented here illustrate the value ofa multidisciplinary approach to the characterization of very old fossil material and validate sequence data reported elsewhere ( 10). The inclusion of fossil-derived molecular sequences into existing phytogenies may provide greater resolution and may allow reconstruction of character evolution beyond what is currently possible. Elucidating modifications to ancient molecules may shed light on patterns of degradation and diagenesis. The presence of original molecular components is not predicted for fossils older than a million years ( 1 - 7), and the discovery of collagen in this wellpreserved dinosaur supports the use of actualistic conditions to formulate molecular degradation rates and models, rather than relying on theoretical or experimental extrapolations derived from conditions that do not occur in nature.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |