
Chiroptera BLUMENBACH, 1779
|
publication ID |
https://doi.org/10.2478/if-2017-0021 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF8797-FFD7-FFA8-FF7F-FF71BA62FDF2 |
|
treatment provided by |
Diego |
|
scientific name |
Chiroptera BLUMENBACH, 1779 |
| status |
|
Order Chiroptera BLUMENBACH, 1779 View in CoL
Bats are relatively common and taxonomically diverse in European fossil sites from Early and Middle Miocene ( Storch 1999, Rosina and Rummel 2012). The geologically oldest bat record for the Caucasus was reported from the early Middle Miocene fauna of Belomechetskaya, Scotopilus sp. and Tadarida sp. ( Gabunia and Bendukidze 1984). But for the Late Miocene only a low number of fossil sites with bat remains are known and the diversity of the bats is limited. For example, only about 5 – 6 species representing modern Vespertilionid genera are known from the early and middle Turolian of southern Ukraine ( Rosina and Sinitsa 2014); the early Turolian (MN 11) fauna of Eichkogel in Austria documents the single species, Paleptesicus cf. noctuloides ( Zeigler 2006) . In Russia, the only described record of a Late Miocene bat, identified as Vespertilio cf. villanyiensis, comes from the fauna of Morskaya 2 (late Turolian, MN 13) ( Rossina et al. 2006).
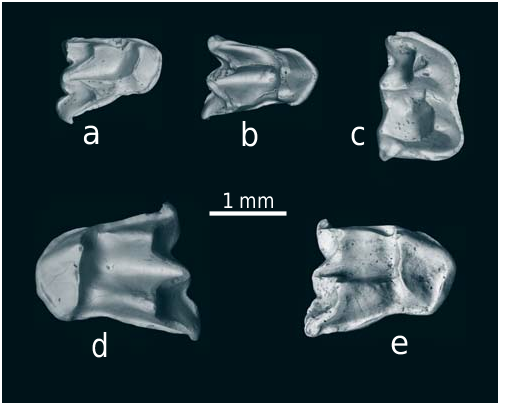
Our new bat material originated from two distinct sites, Volchaya Balka and Gaverdovsky. Both sites are nonkarstic, which in general rarely contain bat remains ( Sigé and Legendre 1983, Rosina et al. 2015). Bats are represented by five isolated cheek teeth: one left M1 or M2, three right M2 and one right m1 or m2. All specimens have partly damaged roots, but the cusps are preserved quite well; only one specimen ( Text-fig. 9a View Text-fig ) has a partly broken parastyle.
For identification of material we compared it with museum specimens of modern bats from different families and genera, taken from the collection of Moscow Zoological Museum (ZMMU). List of species used for comparison includes: Rhinolophus fumigatus , Hipposideros abae , Chaerephon pumila , Ch. nigeriae , Miniopterus schreibersi , Kerivoula kachinensis , Myotis alcathoe , M. annectans , M. blythii , M. dasycneme , M. daubentonii , M. indochinensis , M. montivagus , M. muricola , M. mystacinus , M. nattereri , M. welwitchii , Plecotus auritus , P. turkmenicus , Pipistrellus kuhlii , Nyctalus noctula , N. leisleri , Vespertilio murinus , V. sinensis , Eptesicus serotinus , E. nilssonii , Hesperoptenus tickelli , Arielulus circumdatus , and Scotophilus kuhlii . First of all, from that comparison we came to conclusion that all the five specimens represent the family Vespertilionidae .
The specimen in Text-fig. 9d View Text-fig (left M1 or M2) from Volchaya Balka in morphology and proportions (shape of protocone, stage of hypocone reduction, almost open trigon basin with deep fossa, proportionally unwidened stylar shelf) is mostly similar to large serotines of the genus Eptesicus (especially with M2 of E. serotinus ). In size (L = 1.85, W = 1.95 mm), this molar also corresponds to E. serotinus . According to direct comparison, in proportions the fossil molar somewhat differs from the teeth of Vespertilio and small serotines, such as E. nilssonii ; other large nycticeiines, such as Arielulus and Hesperoptenus , have different proportions and shape of the protocone and a more shallow fossa in the trigon basin. The Late Miocene fauna of Eichkogel in Austria (Ziegler 2006), similar in age to the faunas under study, yielded the single nycticeiine, Miostrellus noctuloides . This form, as well as other Miostrellus , is definitely smaller than E. serotinus ( Rosina and Rummel 2012) and our specimen. Another Late Miocene European serotine, E. campanensis , is similar to modern E. serotinus in both size and proportions. This fossil species was described from France and later found in several sites across Europe ( Rosina and Sinitsa 2014). Eptesicus kowalskii , which occurred sympatrically and synchronously, has a similar morphology but is somewhat smaller (ibid.).
Other upper teeth we identified as belonging to Myotis spp. The only lower molar ( Text-fig. 9c View Text-fig ) does not show features which exclude its affiliation to Myotis . According to its size, it may represent the same species as the upper molar in Text-fig. 9e View Text-fig , found in the same locality (Gaverdovsky). Smaller Myotis teeth, Text-fig. 9a and 9b View Text-fig from Volchaya Balka, are similar in size, shape and morphology and likely belong to the same species. All three upper molars tentatively assigned to Myotis have relatively well developed hypocones and postprotocrysts, and lack paraconules. We did not find this combination in European myotines, but it occurs in some South-East Asian bats from muricola and montivagus species groups. The dimensions of the teeth ( Text-fig. 9a, b View Text-fig ) are L = 1.1 and 1.15, W = 1.4 and 1.5 mm, respectively. In size they are similar to smaller modern Myotis , in particular to M. muricola . They are morphologically specific in the more concave posterior edge. The cingulum is almost interrupted in this concavity while in all inspected modern Myotis there is no cingulum break or even a significant constriction in the same place. Specimens in Text-fig. 9e View Text-fig and, probably, Text-fig. 9c View Text-fig from Gaverdovsky belong to a definitely larger bat with the upper molar size, L = 1.6, W = 1.85, and the lower molar, L = 1.7, Wtrd = 1.0, Wtld = 1.15 mm. This is comparable to larger members of the montivagus species group (almost identical in size to cheek teeth of M. indochinensis ), with which the specimen Text-fig. 9e View Text-fig has a certain morphological similarity. Of the European fossil mouse-eared bats, M. podlesicensis from the Early Pliocene of Poland is most similar in size ( Ziegler 2003, Rosina and Kruskop 2011).
All the available teeth were found in deposits of a shallow water basin. It is known that E. campanensis was described from similar deposits ( Bonis et al. 1999, Rosina and Sinitsa 2014). The foraging activity of many modern Myotis is also closely linked to water pools or directly with the water surface (e.g., Findley 1993). It is thus possible that a similar behavior could have been inherent in the fossil species. Judging from the absence of wear signs and the very sharply pronounced tips and crests on all five teeth, they belong to young individuals, which became independent only shortly before fossilisation. This may imply that both Late Miocene burials at Volchaya Balka and Gaverdovsky were situated near sites favourable for roosting and reproduction of insectivorous bats.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |