
Laimoloima, Gustafsson & Adam & Zou, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5087.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:0CE51AC4-E75F-43DE-B9F5-56247B75F394 |
|
DOI |
https://doi.org/10.5281/zenodo.5827925 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB87B6-DF6A-EE09-29F7-24FFFC83FF7E |
|
treatment provided by |
Plazi |
|
scientific name |
Laimoloima |
| status |
gen. nov. |
Laimoloima new genus
Type species: Laimoloima tandani n. sp.
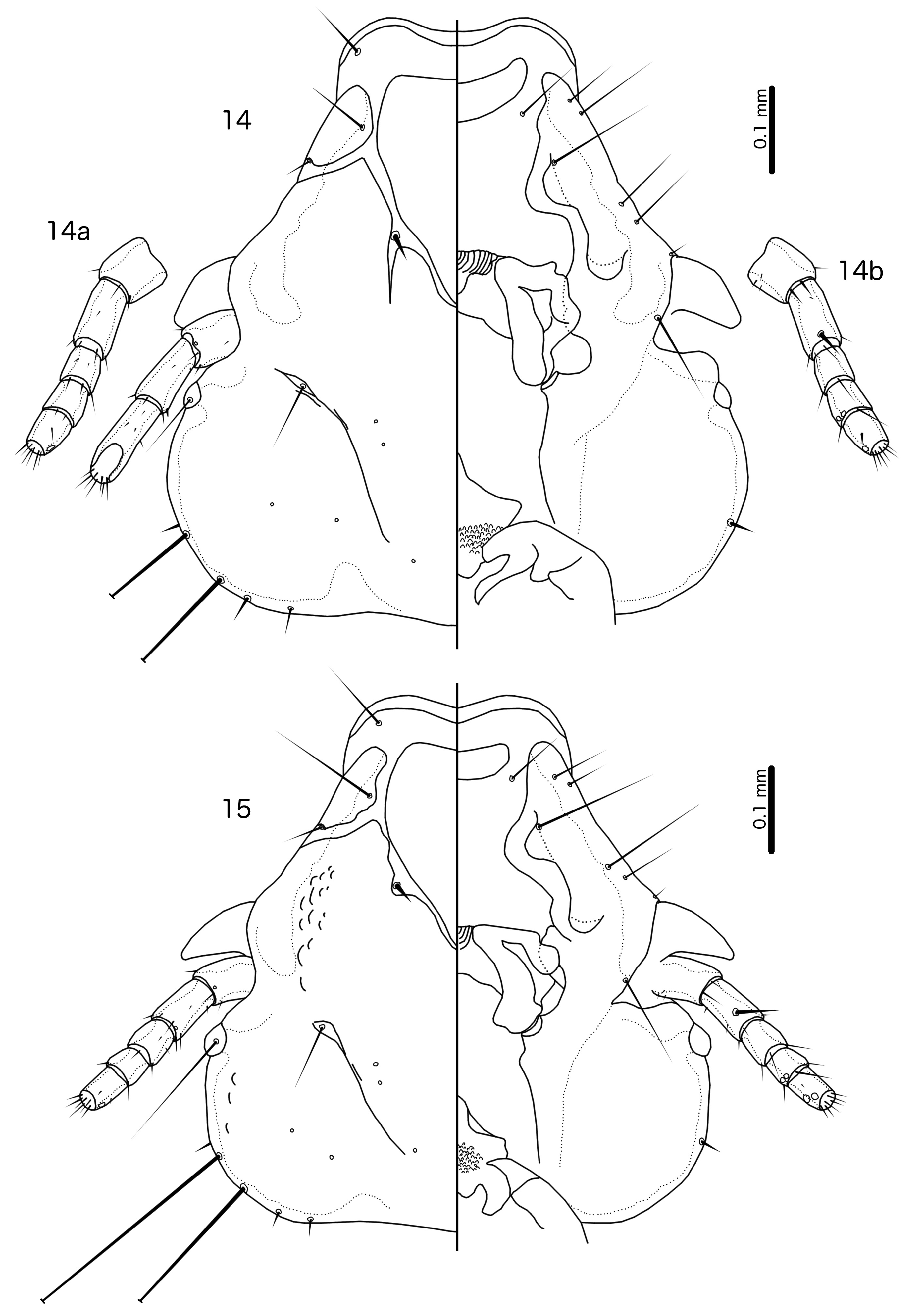
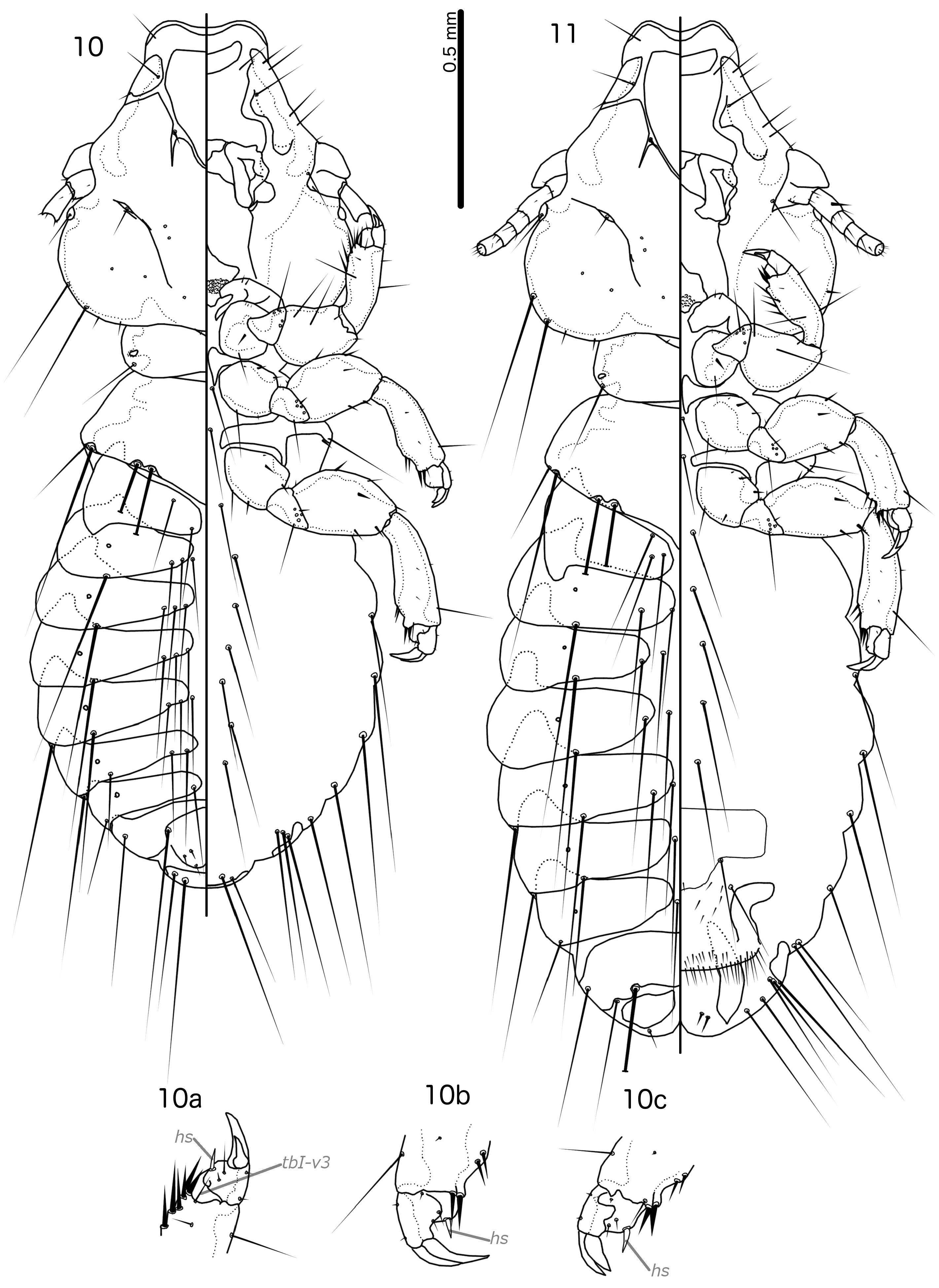
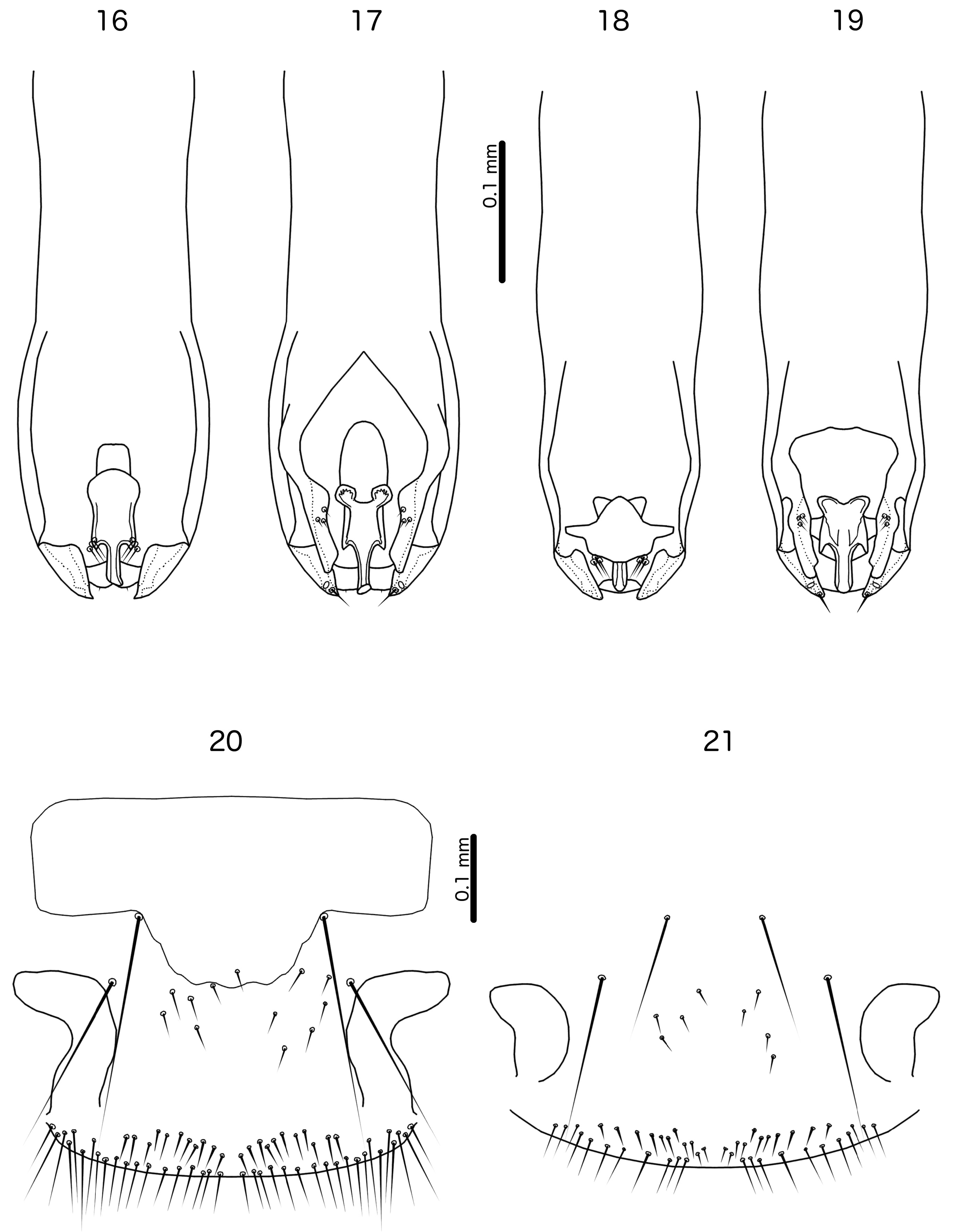
Diagnosis. Laimoloima is morphologically closest to Penenirmus . For comparison, refer to figures 4 and 5 in Sychra et al. (2014) to see the diagnostic characters of Penenirmus against those of Laimoloima as enumerated and illustrated in this paper. These two genera can be separated by the following characters: (1) head hyaline margin with marginal sclerotisation in Laimoloima ( Fig. 14 View FIGURES 14–15 ), but without sclerotisation in Penenirmus ; (2) hyaline margin arising laterally near as2 and thus extending lateral to sclerotised part of frons in Laimoloima ( Fig. 14 View FIGURES 14–15 ), but restricted to median part of frons in Penenirmus ; (3) marginal carina clearly interrupted laterally at point where dorsal preantennal suture reaches head margin in Penenirmus , but not interrupted at that point in Laimoloima ( Fig. 14 View FIGURES 14–15 ); (4) head seta as3 clearly dorsal, situated on hyaline margin in Laimoloima ( Fig. 14 View FIGURES 14–15 ), but marginal and situated on sclerotised part of head in Penenirmus ; (5) head seta as1 clearly dorsal in Laimoloima ( Fig. 14 View FIGURES 14–15 ), but marginal in Penenirmus ; (6) head seta mts1 short in Laimoloima , and in males clearly situated ventrally [in P. rafflesi ( Dalgleish, 1967) this seta appears to be situated near the eye, but we have not seen any specimens to confirm this character], but mts1 is a macroseta in Penenirmus ; (7) head seta mts2 is a macroseta in Laimoloima but is short in Penenirmus ; (8) tergopleurites II–VIII medianly separated in both sexes in Laimoloima ( Figs 10–11 View FIGURES 10–11 ), but medianly continuous (may be indented anteriorly) in Penenirmus ; (9) ventral side of abdomen of both sexes with only one sts on each side in Laimoloima ( Figs 10–11 View FIGURES 10–11 ), but with multiple sts on each side in Penenirmus ; (10) ventral endomere of male genitalia with posterior and anterior lobes in Laimoloima ( Fig. 19 View FIGURES 16–21 ), but without lobes in Penenirmus .
Description. Both sexes. Head rounded, trapezoidal ( Fig. 14 View FIGURES 14–15 ), preantennal area broad, relatively short, typically with concave frons. Hyaline margin broad, arising laterally near site of as2, and with clear sclerotisation of margin. Marginal carina interrupted medianly but not laterally. Dorsal preantennal suture completely surrounds dorsal anterior plate; in all specimens examined, the suture extends slightly distally posterior to ads, as illustrated in Fig. 14 View FIGURES 14–15 , but it is not clear if this is an artifact of mounting. Ventral carina interrupted medianly, extended anteriorly but does not reach frons. Head chaetotaxy as in Fig. 14 View FIGURES 14–15 ; as1 and as3 clearly dorsal; pas and pos absent (pos may be present in B. rafflesi ); avs3 displaced anterior from bend in ventral carina; mts1 clearly ventral at least in males, short seta; mts2–3 macrosetae; s5–7 absent. Dorsal postantennal suture present, with pns situated in suture but s3–4 situated medial to suture. Gular plate present, central part decorated. Thoracic and abdominal segments as in Figs 10–11 View FIGURES 10–11 . Claws of all tarsi of dissimilar size ( Figs 10a–c View FIGURES 10–11 ), and segment I of all tarsi with small, flattened hyaline seta on median margin (hs in Figs 10a–c View FIGURES 10–11 ). Leg setae cI-v3, tI-v2, fI-V4, fI-p1, fII-a2, fII-a5, fIII-a2, fIII-a5 absent; fII-d1 and fIII-d1 displaced toward centre of femur; cI-v1, tbI-d1–5, tbI-dm1, tbII-p1–3, tbII-dm3, tbII-v2, tbIII-p1–3 long and spine-like; tbI-v1 very long; tbI-a1 longer than leg seta tbI-a2; long and slender setae here called tbI-v3 ( Fig. 10a View FIGURES 10–11 ) present medial to tbI-spn on ventral side; trochanter II–III each with two microsetae near anterior margin (tII-a1–2 and tIII-a1–2, respectively). Tergopleurites II–VIII medianly separated in both sexes. Tergopleurite II extended into postero-lateral hook. Sternal and subgenital plates lightly sclerotised in all specimens examined, and may be absent (not illustrated).
Male. Basal apodeme long, tongue-like ( Fig. 16 View FIGURES 16–21 ), but anterior end diffuse in specimens examined. Parameres fused to basal apodeme, pst1 sensilla, pst2 microsetae, in some species very stout. Endomere elongated, with distinct lobes on both ends: distal lobes pointed, and associated with three setae each side dorsally; proximal lobes on ventral side rounded. Distal endomere elongated. Lateral to endomere are paired elongated plates of unknown derivation, each with three small setae at about mid-length; anteriorly these plates appear to be fused to basal apodeme, but this is not clear in specimens examined. Hyaline section of genitalia distal to endomere poorly visible, but one minute seta visible on the distal margin on each side; lateral extent of this hyaline section is not visible in examined specimens, and only the distal margin is illustrated.
Female. Vulval margin with numerous long, slender marginal setae and numerous long, stout submarginal setae; short, slender setae scattered on lightly sclerotised subgenital plate.
Host distribution. Piciformes : Megalaimidae .
Geographical range. Indo-Malayan region.
Etymology. The name Laimoloima is constructed from “ laimós ”, Greek for “throat”, referring to the family name of the host ( Megalaimidae ), and “ loimós ”, Greek for “pest”. Gender: feminine.
Remarks. We include in Laimoloima two new species, described below, and three previously described species. We have not seen any specimens of Docophorus limbatus Piaget, 1885 (as Penenirmus limbatus in Hopkins & Clay 1952: 275) from Psilopogon corvinus (Temminck, 1831) , and the original illustration is poor. Piaget (1885: 4) states that the frons has a “coloured border”, which we interpret as referring to the thin band of sclerotisation of the hyaline margin. Therefore, we tentatively include Docophorus limbatus Piaget, 1885 in Laimoloima . Furthermore, Penenirmus rafflesi Dalgleish, 1967 and Penenirmus zeylanica Dalgleish, 1967 are also included in Laimoloima , based on the original descriptions which, as far as it is possible to see, share all the characters of the new genus.
In Tendeiro’s (1961) illustrations of Penenirmus guineensis , the frons appears to be sclerotised, the dorsal preantennal suture is present, and as3 is illustrated on the dorsal side. Both, Tendeiro’s (1961) photo of P. bidentatus with a sclerotised hyaline margin and his photo of the male genitalia of P. guineensis suggest a relationship with Laimoloima . However, in the phylogeny of Johnson et al. (2021: fig 1), P. guineensis is embedded in a clade with lice from other African barbets and a woodpecker, sister to a clade of Penenirmus from Passeriformes , and both clades together are sisters to all other Penenirmus sensu lato and Picophilopterus . Nevertheless, as we have not examined any specimens of any African species, we retain P. guineensis and P. bidentatus in Penenirmus until they can be redescribed.
In the phylogeny of Johnson et al. (2021), one louse belonging to the Penenirmus -complex from Psilopogon chrysopogon (Temminck, 1824) (Megalaimidae) was placed as sister to a large clade containing several species of Picophilopterus , as well as lice from honeyguides ( Indicatoridae ), Neotropical barbets ( Capitonidae ) and two African barbets ( Lybiidae ). Presumably, the specimen from Psilopogon chrysopogon belongs to Laimoloima , but its morphology is not known to confirm its taxonomic position. Although the phylogeny of Johnson et al. (2021) contains only one species from Asian barbets, its position would indicate that the lice of the Penenirmus -complex from these birds are not closely related to those from African and Neotropical barbets, and that some morphological similarities between species of Laimoloima species and those from African barbets (e.g. the sclerotised central part of the hyaline margin) may be the result of convergent evolution.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.