
Trechisibus (Trechisibiodes) apukhapiensis, Ruiz-Tapiador & Delgado, 2016
|
publication ID |
https://doi.org/10.5281/zenodo.10905236 |
|
DOI |
https://doi.org/10.5281/zenodo.10948923 |
|
persistent identifier |
https://treatment.plazi.org/id/03A8D771-D90E-FFD3-5B3E-82C76D762158 |
|
treatment provided by |
Felipe |
|
scientific name |
Trechisibus (Trechisibiodes) apukhapiensis |
| status |
new species |
Trechisibus (Trechisibiodes) apukhapiensis View in CoL , new species
Holotype. Male labeled/ Perś, Puno, montańa Apu Khapia 68º 08'57'’ W, 16º 20' 41'’ S, 4 462 m, 19-IX-2012, P. Delgado leg. deposited in National Museum of Natural History in Lima ( MNHNL). GoogleMaps
Type locality. Perś, Puno Region, Chucuito Province, Pomata District , Apu Khapia Mountain 68ŗ 08' 57'’ W, 16ŗ 20' 41'’ S, 4 462 m.
Paratypes (18). Four males with same data as holotype, deposited col. P. Delgado (2) and col. I. Ruiz-Tapiador (2); One male Perś, Puno, montańa Apu Khapia 68º 08' 57' ‘W, 16º 20’ 41'’ S, 4 462 m, 06-IX-2012, P. Delgado leg. deposited in MNHNL (1) ; two female, Perś, Puno, montańa Apu Khapia 68º 08' 57'’ W, 16º 20' 41'’ S, 4 462 m, 06-IX-2012, P. Delgado leg. deposited col. P. Delgado (1) y col. I. Ruiz-Tapiador (1). Seven males Perś, Puno, montańa Apu Khapia 68º 08' 57'’ W, 16º 20' 41'’ S, 4 462 m, 20-XII-2012, P. Delgado leg. deposited in MNHNL (2), col P. Delgado (2) and col I. Ruiz-Tapiador (2); five females, Perś, Puno, montańa Apu Khapia 68º 08' 57'’ W, 16º 20' 41'’ S, 4 462 m, 20-XII-2012, P. Delgado leg. deposited col. MNHNL (2), P. Delgado (2) and I. Ruiz-Tapiador (1) GoogleMaps .
Etymology. The new species is named after the type locality, the Apu Khapia Mountain.
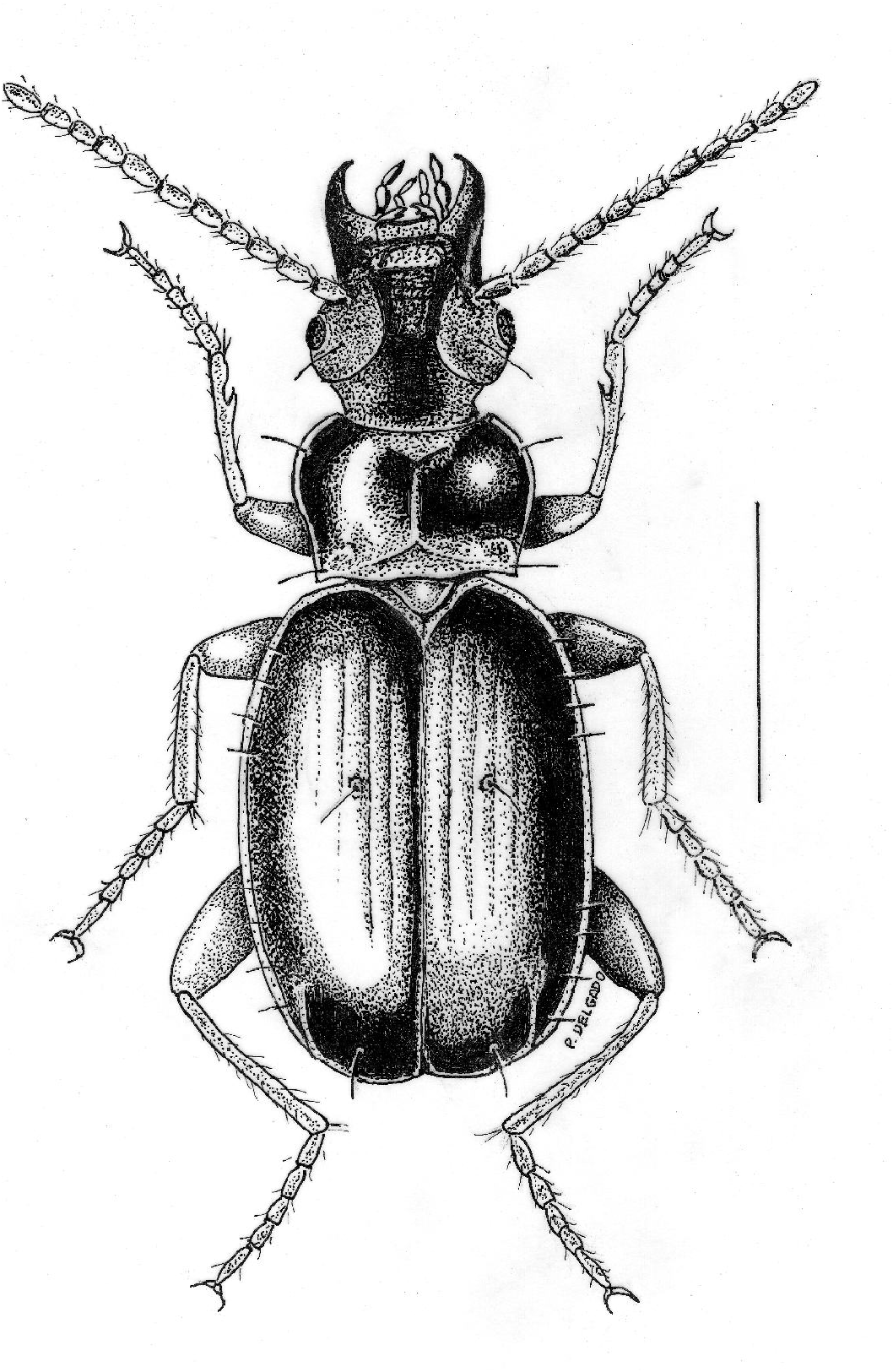
Description. Small for the genus ( ABL range 2.9– 3.2 mm, mean 3.04 ± 0.09 mm, n=19). Sexes of approximately equal size: ABL range of males 2.9– 3.2 mm, mean 3.04 ± 0.10 mm, n=12; ABL range of females 3.0– 3.1 mm, mean 3.05 ± 0.07 mm, n=7. Habitus ( Figure 1 View Fig ) markedly convex, ovoid ( WE / ABL 0.35 ± 0.01), head normally proportioned for the genus (WH/WPm 0.82 ± 0.01), pronotum narrow compared to elytra (WPm/ WE 0.73 ± 0.04). Body color ranges from reddish brown to dark brown, appendages much lighter, testaceous. Micro-sculpture fine but distinct across dorsal surface of pronotum and most of head. On head only the vertex lacks micro-sculpture. Elytra with well-developed polygonal micro-sculpture.
Pronotum moderately convex and comparatively strongly transverse (WPm/ LP 1.46 ± 0.19), with straight margins, moderately constricted at posterior region (WPm/WPp 1.35 ± 0.06). Anterior angles evident and slightly prominent. Posterior angles slightly obtuse (100–105°). The width between posterior angles is greater than between anterior angles (WPa/WPp 0.82 ± 0.06).
Elytra markedly convex, slightly depressed along suture, of normal length for genus ( LE / ABL 0.53 ± 0.01). Micro-sculpture consisting of polygonal meshes, distinct on the whole elytral surface, which is slightly shiny. Shoulders rounded, with angulose humeral border ending at elytral base near the 4th stria. Lateral border of elytra wide and sharply narrowed backwards ending in a slight preapical emargination. Elytral tip broad and rounded. Second to seventh stria vestigial; first stria shallow but evident on the whole length; eighth stria deeply impressed on the whole length; apical recurrent striole short, sub-rectilinear, ending at level of seventh stria and with a remarkable apical carina. Chaetotaxy: juxtascutellar pore present; one setiferous dorsal pore on the third stria, at basal fifth and a little behind middle. Preapical pore placed backwards and closer to recurrent striole than to suture.
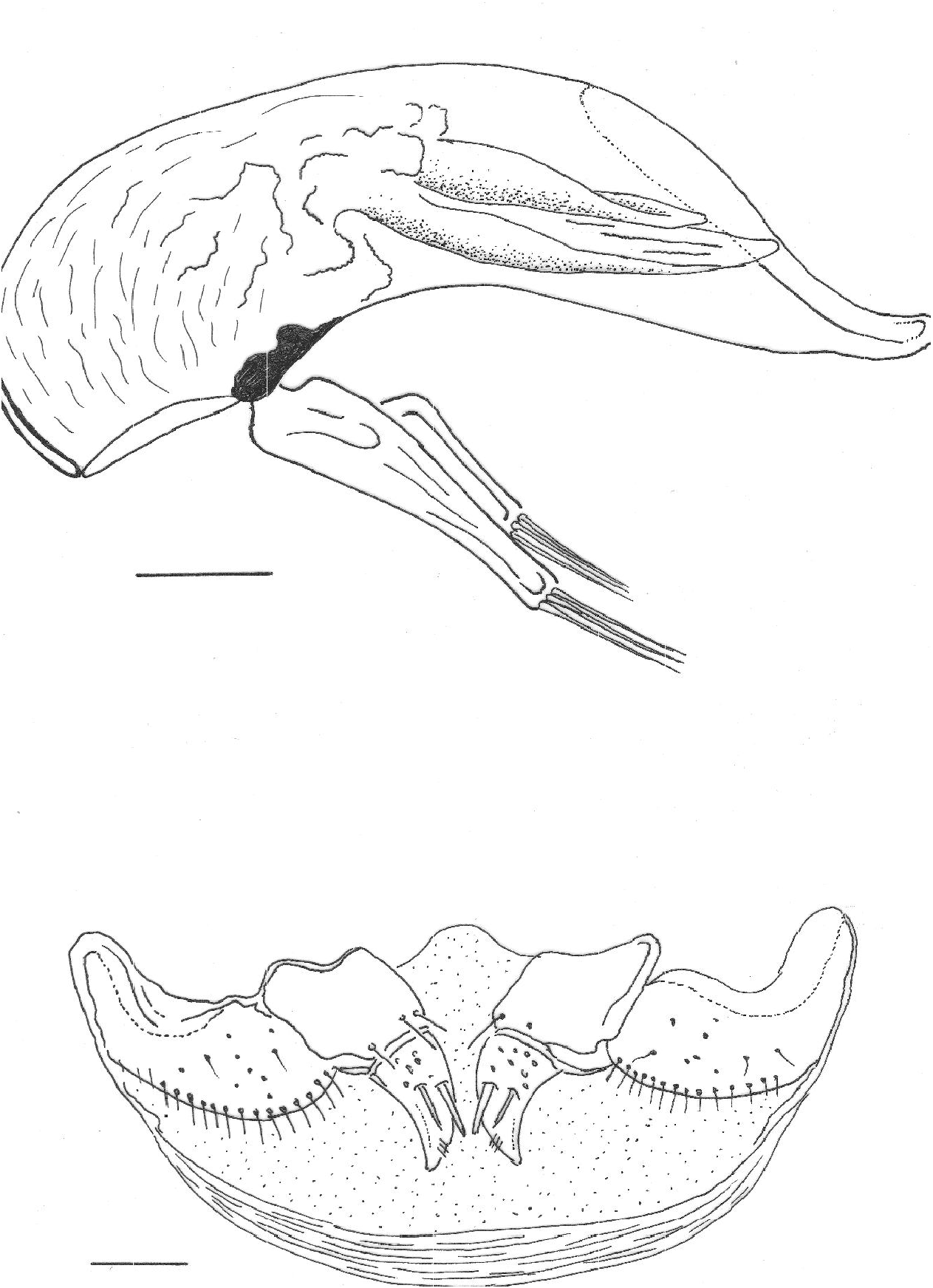
Aedeagus ( Figure 2 View Fig ) moderately sclerotized, small (length 0.70 mm), slender, with basal bulb enlarged; median lobe, in lateral view, regularly tapering from base to apex, little and regularly curved; apex long, relatively slender and strongly bent downwards. Inner sac with a copulatory piece in the middle part shaped as a long open gutter, bordered by small striated elements. Parameres narrow and long, reaching the apical fourth of the median lobe, each provided with four apical setae.
Distribution. At present T. apukhapiensis sp. n. is only known from the type locatity, Apu Khapia Mountain.
Habitat. Beetles were collected under stones laying on rather dry soil, near a lagoon at 4 620 m. a.s.l.n. At this site T. apukhapiensis sp. n. was collected in sintopy with other Trechinae.
Differential diagnosis. The new species is included in the subgenus Trechisibiodes , based on the presence of only one discal seta on the elytron and on the relative position of the apical seta ( Uéno 1972). As stated in the introduction, another 14 species of this subgenus are known to occur in the study area, from which the new species can be clearly differentiated by the features of male and female genitalia, in combination with some external morphological characters.
T. apukhapiensis sp. n. is easily distinguishable from a first group of species composed of Trechisibus ( Trechisibiodes) veneroi Etonti & Mateu, 1992 ; T. cuzcoensis Etonti & Mateu, 1996 ; T. nicki Schweiger, 1959 ; T. franzi Mateu & Negre, 1972 and T. peruvianus Jeannel, 1927 . In this group, as can be observed ( Etonti & Mateu 1992, 1996, Jeannel 1927, Mateu & Negre 1972, Schweiger 1959), the apex of the aedeagus, in lateral view, is curved ventrally, as well as the well developed sagital flap at the base of the basal bulb in all these species. In T. apukhapiensis sp. n., however, the apex is virtually horizontal and the sagital flap is not developed, and only observable as a small, slightly more chitinized area in an homologous position.
Within a second group of species of Trechisibiodes , T. apukhapiensis sp. n. resembles several species, except for the presence in those species of the sagital flap at the base of the bulb, a medial lobe (in lateral view) curved dorsally and ending in a hook. Both characters allow differentiating without problems of T. apukhapiensis sp. n. from T. laresensis Etonti & Mateu, 1996 ; T. theresiae Etonti & Mateu, 1996 ; T. schmidti Uéno, 1971 and T. wardi Etonti, 2003 ( Etonti 2003, Etonti & Mateu 1996, Uéno 1971).
Among the geographically closest species with known male genitalia, T. bohorquezae Etonti & Mateu, 1992 and T. orophilus Mateu & Etonti, 2002 are those with an apex more akin to that of T. apukhapiensis sp. n. ( Etonti & Mateu, 1992, Mateu & Etonti 2002).
From T. bohorquezae , it may be distinguished by the general aspect of the medial lobe of the aedeagus and the narrow, long and sagital flap at the base of the bulb in T. bohorquezae ( Etonti & Mateu 1992) , as well as by a combination of traits absent in T. apukhapiensis sp. n.
T. orophilus is probably the species with an aedeagus more akin to that of T. apukhapiensis sp. n.., however, in the case of the latter species the medial lobe of the aedeagus, in lateral view, is strongly bent ventrally, the apex is slightly narrowed and the morphology of the chitinized piece from the internal armature is different ( Mateu & Etonti 2002). Additionally, according to the original description of T. orophilus , the basal foveae of pronotum are deep, while in T. apukhapiensis n. sp. they are very shallow.
There are three species of Trechisibiodes in the study area with unknown male genitalia: T. ukupachensis Trezzi, 2007 ; T. gigas Trezzi, 2007 and T. pygmaeus Jeannel, 1930 . In the case of the first two, the morphology of the external female genitalia is characterized in the original descriptions ( Trezzi 2007), but not in the latter ( Jeannel 1930).
Among the characters separating T. ukupachensis from T. apukhapiensis sp. n., what stands out is the dorsal surface of the gonocoxites in the former, which has only one spine, while in the new species two well developed spines are clearly visible. Moreover, T. ukupachensis is larger, with 4.8 mm length, while T. apukhapiensis sp. n., with 3.01 mm length, thus being one of the smallest species of the subgenus.
T. gigas , is a peculiar species within the subgenus by its exceptionally large size, which is more than twice the length of T. apukhapiensis sp. n. But it also differs in the lesser robustness of the spines of the gonocoxite and in the relative position of both spines which in T. gigas are stepped, while in T. apukhapiensis sp. n., these are practically in parallel.
The difference with T. pygmaeus is more problematic, given that the original description is quite succinct and only based on characters associated to the external morphology of the single female holotype. Fortunately both species can be well distinguished since in T. apukhapiensis sp. n. the tibiae present a groove in their external face, while in T. pygmaeus the description explicitely refers to the lack of this trait.
Biogeographical notes. The discovery of the new species of Trechinae is significant due to the biogeographical position, filling a gap in the knowledge of the distribution of this genus in the south of the Peruvian Andes (Figure 3).
| MNHNL |
MNHNL |
| ABL |
Adviesbureau voor Bryologie en Lichenologie |
| LP |
Laboratory of Palaeontology |
| LE |
Servico de Microbiologia e Imunologia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Trechinae |
|
Genus |