
Eudendrium bleakneyi, Calder, Dale R., 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.556851 |
|
publication LSID |
lsid:zoobank.org:pub:985C0239-D00C-457D-B593-76A3081BCEEA |
|
DOI |
https://doi.org/10.5281/zenodo.6015990 |
|
persistent identifier |
https://treatment.plazi.org/id/03A787C7-4931-FFA2-FF58-F9B0FAB0F9DB |
|
treatment provided by |
Plazi |
|
scientific name |
Eudendrium bleakneyi |
| status |
sp. nov. |
Eudendrium bleakneyi View in CoL , sp. nov.
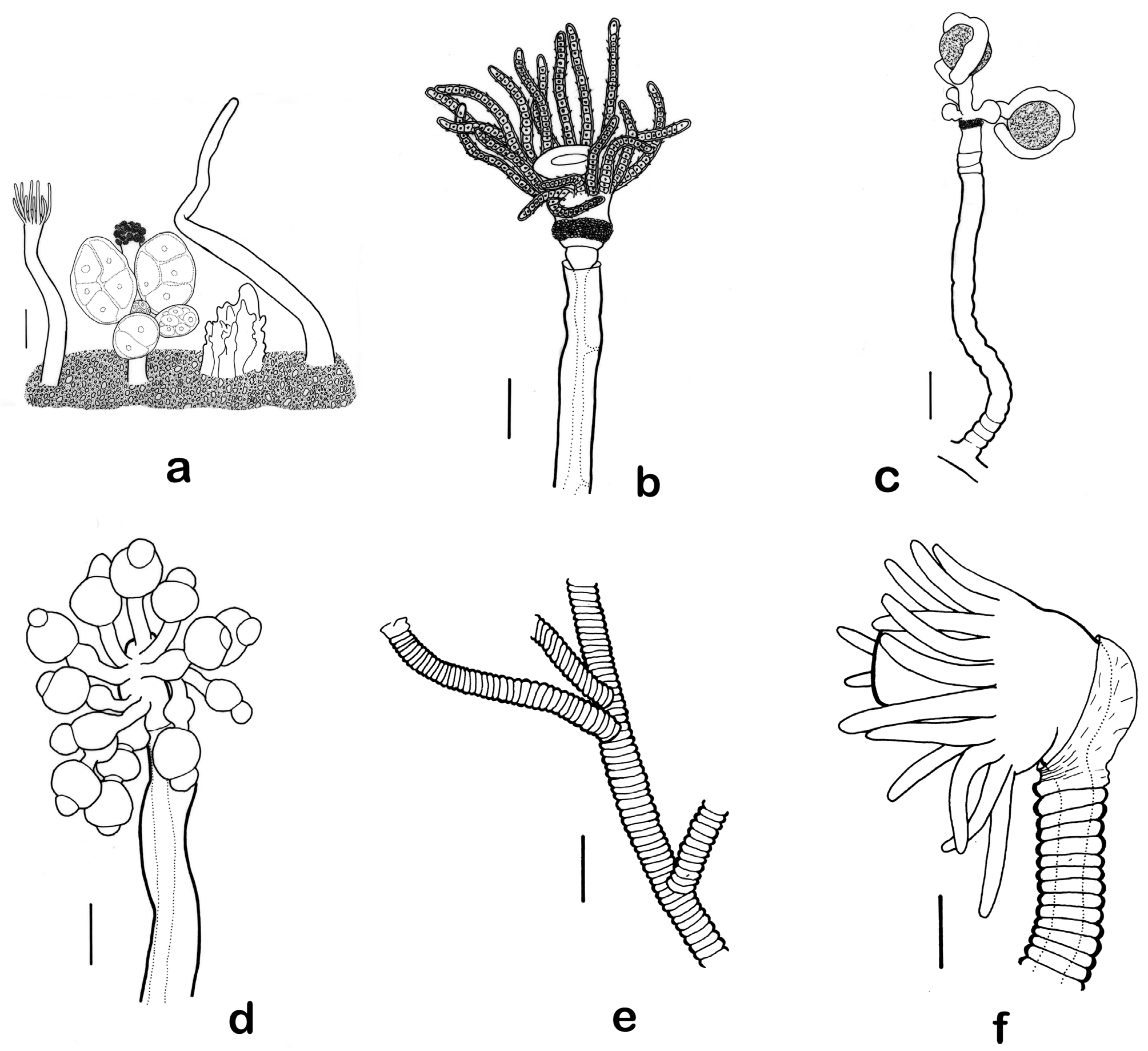
Figs. 13 View FIGURE 13 b–d, 14, 15
Material examined. HOLOTYPE. NB: St. Croix River, St. Andrews, off Joe's Point , 45°04’30”N, 67°05’20”W, 16 m, scallop drag, 06.ix.1984, several colony fragments, up to 11 mm high, with male gonophores, coll. D. Calder, ROMIZ B1473. GoogleMaps
PARATYPES. NB: St. Croix River, St. Andrews, off Joe's Point , 45°04’30”N, 67°05’20”W, 16 m, scallop drag, 06.ix.1984, one colony, 1.1 mm high, with single blastostyle having two female gonophores, coll. D. Calder, ROMIZ B1381. GoogleMaps
NB: St. Croix River, St. Andrews, off Joe's Point , 45°04’30”N, 67°05’20”W, 16 m, scallop drag, 06.ix.1984, one colony, 6 mm high, one blastostyle with male gonophores, coll. D. Calder, ROMIZ B1472. GoogleMaps
NB: St. Croix River, St. Andrews, off Joe's Point , 45°04’30”N, 67°05’20”W, 16 m, scallop drag, 06.ix.1984, one colony, 5.5 mm high, without gonophores, coll. D. Calder, ROMIZ B1475. GoogleMaps
Etymology. The specific name honours Dr. J.S. Bleakney of Acadia University, an esteemed mentor who contributed greatly to the content of this work.
Description. All parts of hydroid exceptionally minute. Colonies mostly erect, arising from a creeping hydrorhiza, reaching 11 mm high, occasional parts of colony bearing single pedicels. Hydrocaulus monosiphonic, contorted, sparsely and irregularly branched, about 0.05–0.09 mm in diameter, with branches resembling hydrocaulus. Perisarc thin, amber-coloured in older parts, clear in younger parts, covering all parts of colony except hydranth distal to perisarc groove, annulated at bases of hydrocauli, branches, and ultimate branchlets, with occasional wrinkles elsewhere but mostly smooth. Hydranths cup-shaped to tumbler-shaped, about 0.2 mm long from perisarc groove to hypostome, 0.1 mm wide at level of tentacles, with distinct perisarc groove basally and knob-shaped to flared hypostome distally. Tentacles solid, filiform, in one whorl, about 18–20 in number; tentacular nematocysts arranged obliquely. Base of hydranth immediately above perisarc groove with a variably elevated nematocyst belt containing an extraordinarily dense aggregation of microbasic euryteles. Nematophores absent. Nematocysts of tentacles, hypostome, and nematocyst belt comprising microbasic euryteles only.
Colonies gonochoristic; gonophores fixed sporosacs arising from reduced hydranths. One tiny female colony with a single reduced hydranth having two gonophores, each gonophore with an unbranched, knobby spadix curving over egg, with knobs containing batteries of nematocysts. Male gonophores occurring in a cluster on reduced hydranths having no tentacles, such blastostyles bearing gonophores having 1–3 round to oval chambers, multiple chambers when present in a linear series, terminal chamber bearing a prominent cap or projection heavily armed with microbasic euryteles.
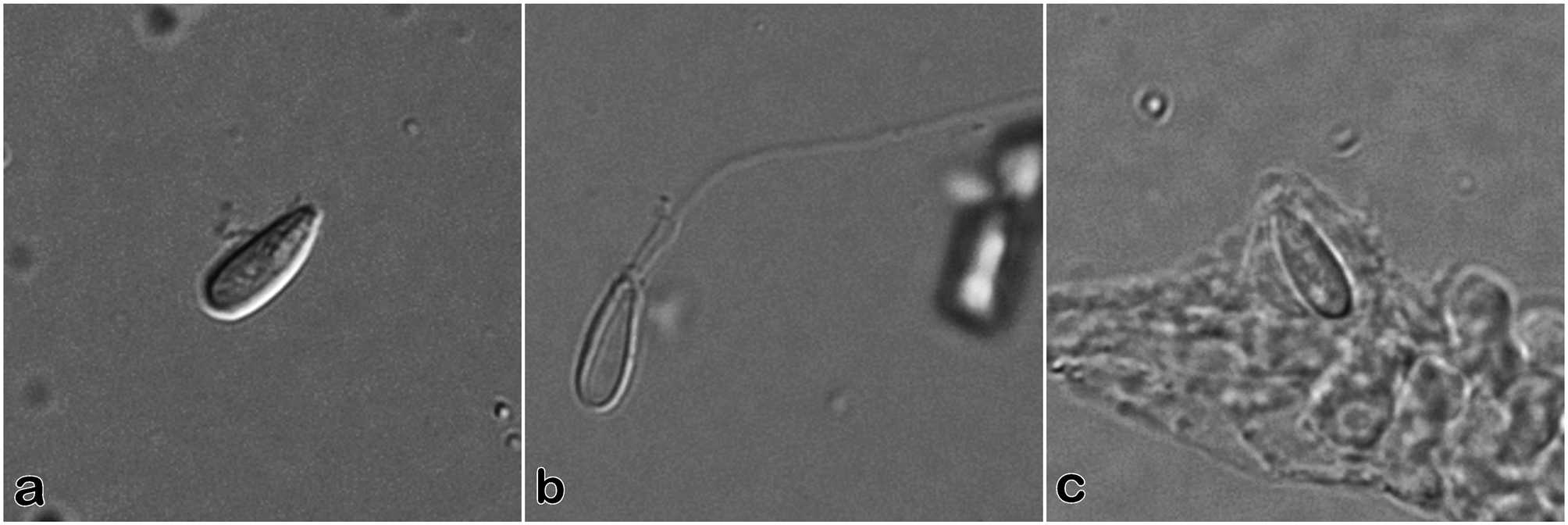
Cnidome ( Fig. 15 View FIGURE 15 )
Hydranths—microbasic euryteles (n = 4): 6.2–8.8 µm long × 2.6–3.1 µm wide (undischarged)
Terminal nematocyst cap, male gonophores—microbasic euryteles (n = 10): 6.6–7.1 µm long × 2.0–2.5 µm wide (undischarged)
Remarks. Species of the anthoathecate hydroid genus Eudendrium Ehrenberg, 1834 are readily distinguished by the characteristic morphology of their hydranths and gonophores. Hydranths are usually large and urn-shaped, with a perisarc groove basally, a single whorl of solid filiform tentacles near the distal end, and a prominent, knobshaped to flaring hypostome beyond. Gonophores arise at least initially from the gastric column of the hydranth, and are fixed sporosacs in all known species of the genus. With certain exceptions, those of the male are arranged in a linear series of bead-like chambers while those of the female consist of a spadix enveloping a single egg. Morphological and molecular evidence both support the hypothesis that the genus is monophyletic ( Marques, Peña Cantero et al. 2000; Moura et al. 2008).
Although the genus Eudendrium is highly distinctive, the numerous nominal species assigned to it can be exceedingly difficult to identify on the basis of morphology alone. Many of them were founded on sterile specimens or on unreliable characters, and are now considered species inquirendae. Characters accorded particular taxonomic importance include the structure of the gonophores, particularly those of the female, the extent of retrogression of reproductive hydranths, and more recently the complement, distribution, shape, and size of nematocysts ( Picard 1951; Watson 1985; Calder 1988, 2010; Marques, Mergner et al. 2000; Marques, Peña Cantero et al. 2000; Puce et al. 2005; Bouillon et al. 2006; Schuchert 2008b). Also perceived to be helpful in the identification of certain species is the presence or absence of buttons or a continuous band of nematocysts around the gastric column of the hydranth, the presence or absence of nematophores, the prominence of the perisarc groove on the hydranth, and the orientation of nematocysts on the tentacles, whether tending to be parallel or oblique to the tentacular axis ( Schuchert 2008b).
As for Eudendrium bleakneyi , sp. nov., its diminutive colony size aligns it with a group of small species included within a so-called “ E. capillare group” (e.g., Watson 1985; Marques, Mergner et al. 2000). That group appears to be morphologically diverse, however, so comparisons were made here with all recognized species of the genus. Particular attention was given to the cnidome, although other morphological characters were considered as well.
Within the genus Eudendrium , 73 nominal species are currently recognized as valid worldwide in WoRMS. With the identity of two of them ( E. attenuatum Allman, 1877 ; E. laxum Allman, 1877 ) being regarded as doubtful (e.g., Puce et al. 2005), 71 species were considered during this study in making comparisons with E. bleakneyi . Of these, 48 are distinctly different from the new species in having “complementary” nematocysts ( Table 2), a designation given to nematocysts in addition to small microbasic euryteles, which are believed to occur in all species of the genus. Therefore, these 48 need not be considered further here in relation to E. bleakneyi .
Of the remaining 23 species, at least 11 resemble E. bleakneyi in having a cnidome comprising only small microbasic euryteles ( E. antarcticum Stechow, 1921b ; E. armatum Tichomiroff, 1887 ; E. capillare Alder, 1856a ; E. corrugatum Watson, 1985 ; E. deciduum Millard, 1957 ; E. deforme Hartlaub, 1905 ; E. japonicum Yamada, 1954 ; E. nambuccense Watson, 1985 ; E. pennycuikae Watson, 1985 ; E. terranovae Watson, 1985 ; E. vervoorti Marques & Migotto, 1998 ). However, all of these species differ from E. bleakneyi in one or more morphological characters. For example, a prominent nematocyst belt, such as that encircling the base of the hydranth in E. bleakneyi , is lacking in all except E. nambuccense and possibly E. antarcticum ( Millard 1975; Watson 1985, 2003; Hirohito 1988; Marques, Mergner et al. 2000; Schuchert 2008b). Moreover, unlike in E. bleakneyi , colonies of E. armatum , E. corrugatum , E. deciduum , and E. terranovae are relatively large and polysiphonic ( Millard 1975; Watson 1985; Schuchert 2008b). Other noteworthy differences from E. bleakneyi include the presence of nematophores in E. armatum and E. japonicum (Hirohito 1988; Schuchert 2008b), the existence of a branched spadix in E. deciduum ( Millard 1957, 1975); the lack of a spadix over the ovum in E. vervoorti ( Marques & Migotto 1998) , and the paired chains of chambers in male gonophores of E. deforme (Watson 2003) . Notably too, some colonies of E. capillare have complementary nematocysts in the form of small isorhizas ( Schuchert 2008b). Eudendrium bleakneyi is very close in morphology to E. nambuccense , differing from the original account of that species ( Watson 1985) in: (1) being sparsely rather than profusely branched, (2) having small microbasic euryteles that are proportionally more slender, (3) having at least some male gonophores with two chambers rather than a single one, (4) having female gonophores on atrophied rather than on partially atrophied hydranths, (5) having hydranths with about 18–20 rather than 24–28 tentacles. Meanwhile, it is unclear from existing descriptions and illustrations whether a nematocyst belt exists in E. antarcticum , although it seems possible (see Marques, Mergner et al. 2000: fig. 9). Nevertheless, that species differs from E. bleakneyi in having female gonophores arising from hydranths that are little if at all reduced ( Stechow 1921b, 1925).
The cnidomes of the remaining 12 species are currently undescribed. Of primary interest in this heterogeneous group is the identity of E. cingulatum Stimpson, 1853 , unreported since its original description from Bay of Fundy at Grand Manan Island, NB. Stimpson’s (1853) account of the species was lacking in detail, and no illustration was provided. Fraser (1918) thought it might be identical with E. annulatum Norman, 1864 , but later simply questioned its validity ( Fraser 1944, 1947a). Earlier, A. Agassiz (1865) had included it, with question, in the synonymy of Bougainvillia superciliaris ( L. Agassiz, 1850) . The hydroid was described by Stimpson as “small,” but no measurements were given. There was no indication that his colonies were fertile, and gonophores of the species remain unknown. The specific name cingulatum , meaning “belted” or “girdled,” is intriguing. While it might refer to a belt of nematocysts on the hydranths, such as that found in E. bleakneyi and certain other species of the genus, Stimpson provided no explanation for his choice of the name, and it is unknown what character the Latin adjective was describing. His species seems to differ from E. bleakneyi in colony form, with branches “…somewhat as in E. rameum …” rather than being sparsely branched or stolonal, and in having pedicels that were strongly annulated, sometimes completely so, rather than being mostly smooth beyond the base. No record of the species was found in a search of online catalogues of the MCZ, NMNH, or YPM, and no type is known to exist. Eudendrium cingulatum is regarded here as a nomen dubium .
None of the other species included in the genus appear from their descriptions to match E. bleakneyi . In the size and habit of their colonies, only two ( E. irregulare Fraser, 1922 ; E. rugosum Fraser, 1940 ) resemble this new species. Those of the remainder ( E. armstrongi Stechow, 1909 ; E. caricum Jäderholm, 1908 ; E. certicaule Fraser, 1938a ; E. cochleatum Allman, 1877 ; E. dispar L. Agassiz, 1862 ; E. distichum S.F. Clarke, 1879 ; E. maldivense Borradaile, 1905 ; E. nodosum Fraser, 1938a ; E. speciosum Fraser, 1945 ) differ in having colonies that are larger and more branched. Other reported differences setting these species apart include (1) the occurrence, on nonaborted hydranths, of male and female gonophores in E. certicaule and E. dispar , and of male gonophores in E. maldivense and E. speciosum ; (2) the presence of oblique annulations at the bases of stems and branches in E. cochleatum ; (3) the existence of nodes at irregular intervals on stems and branches in E. nodosum ( L. Agassiz 1862; Allman 1877; Borradaile 1905; Fraser 1938a, 1945). Finally, stems and branches of E. irregulare differ from those of E. bleakneyi in having occasional wrinkles but no annulations ( Fraser 1922), while those of E. rugosum differ in being strongly wrinkled throughout ( Fraser 1940).
Nine species of Eudendrium have been reported from the Bay of Fundy (see Appendix 1), with two of those ( E. cingulatum Stimpson, 1853 ; E. tenellum Allman, 1877 ) regarded as species inquirendae. Eudendrium bleakneyi , described as new here, is added as a tenth species from the bay.
Microbasic euryteles borne on nematocyst caps of the male gonophores of this species appeared to be slightly smaller and more slender than those of the tentacles and nematocyst ring on the hydranth.
Eudendrium bleakneyi View in CoL was dredged during late summer (06 September 1984) in relatively shallow waters (16 m) from a commercial scallop [( Placopecten magellanicus ( Gmelin, 1791) ] ground in the St. Croix River off St. Andrews, NB. Salinities in that region of the estuary are typically in the polyhaline range (18–30‰), and water temperatures that time of year are normally between 10–15° C (personal observations).
distinguishing microbasic and macrobasic euryteles may be inconsistent from one author to another.
……continued on the next page ……continued on the next page Complementary nematocysts of Eudendrium angustum Warren, 1908 were thought by Millard (1975) to be either macrobasic euryteles or isorhizas, and not microbasic euryteles. Puce et al. (2005) regarded them as large microbasic euryteles.
Cooke (1975) regarded specimens examined by him as questionably referable to Eudendrium breve .
The cnidome of Eudendrium dispar was examined during this study in syntype material from Nahant, Massachusetts (MCZ COEL 34). Complementary nematocysts, in a band around the base of the hydranth, were large microbasic heteronemes, most likely microbasic euryteles.
The collection locality of Eudendrium distichum was said by S.F. Clarke (1879) to be “Ten miles north of Zoblos Island . ” “ Zoblos Island ” was taken to be Holbox Island, Mexico,
Calder (2013: 59).
According to Watson (1985), microbasic euryteles on gonophores of E. nambuccense are slightly larger than those on hydranths.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Hydroidolina |
|
Order |
|
|
Family |
|
|
Genus |
Eudendrium bleakneyi
| Calder, Dale R. 2017 |
E. attenuatum
| Allman 1877 |
E. laxum
| Allman 1877 |
Eudendrium
| Ehrenberg 1834 |
Placopecten magellanicus (
| Gmelin 1791 |