
Halecium praeparvum, Calder, Dale R., 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.556851 |
|
publication LSID |
lsid:zoobank.org:pub:985C0239-D00C-457D-B593-76A3081BCEEA |
|
DOI |
https://doi.org/10.5281/zenodo.6016002 |
|
persistent identifier |
https://treatment.plazi.org/id/03A787C7-4907-FFB4-FF58-FA25FE64FD91 |
|
treatment provided by |
Plazi |
|
scientific name |
Halecium praeparvum |
| status |
sp. nov. |
Halecium praeparvum View in CoL , sp. nov. Figs. 18–20 View FIGURE 18 View FIGURE 19 View FIGURE 20
Material examined. HOLOTYPE
ME: Cobscook Bay , Eastport, near Estes Head, 44°53’36”N, 66°59’37”W, 07.viii.2005, from aquaculture pen, on kelp holdfast, <1 m, one colony, to 0.3 mm high, with gonothecae, coll. D. Calder, ROMIZ B4111. GoogleMaps
Etymology. The specific name is taken from the Latin adjective praeparvum , meaning very small, minute, tiny, in reference to the minuscule size of this hydroid.
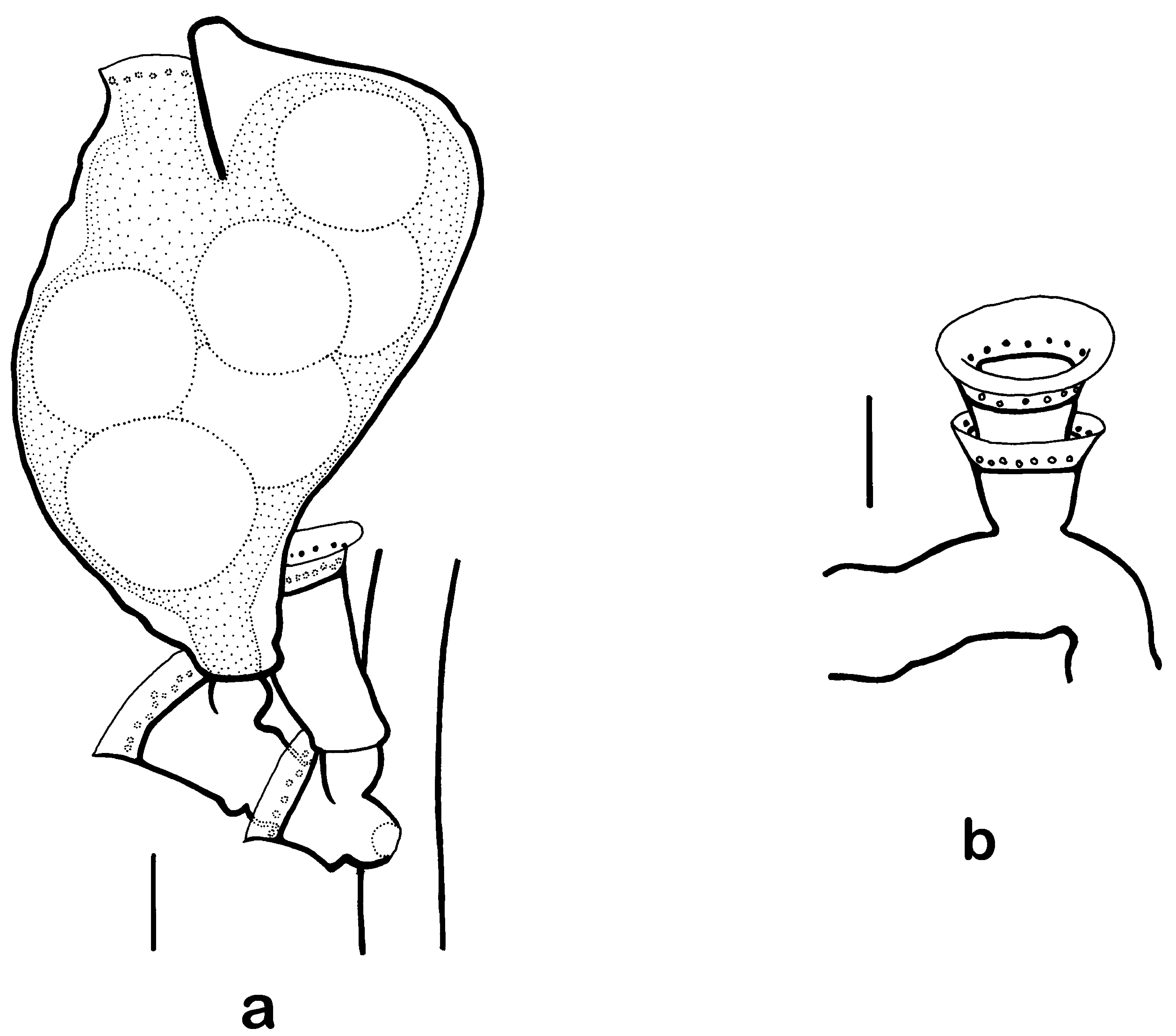
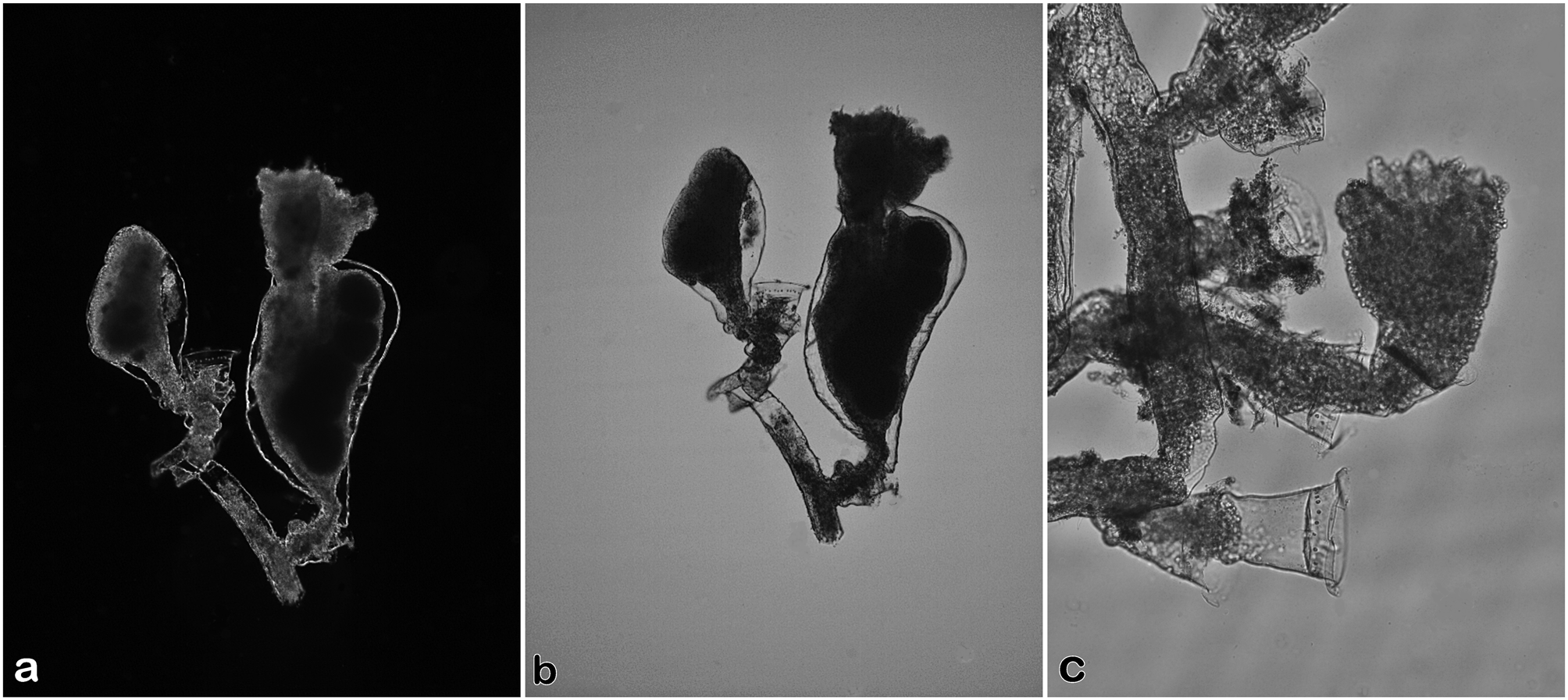
Description. Hydroid colonies minute, stolonal, or erect but with no continuous main hydrocaulus, up to 0.3 mm high, arising from a creeping hydrorhiza that is smooth rather than wrinkled or annulated. Stolonal parts of colony with very short pedicels, occasionally with renovated hydrothecae. Erect colonies comprising no more than three hydrothecae; secondary hydrothecae at the end of a short hydrophore arising from within primary hydrotheca, hydrophore with a basal bulge; any side branches bearing hydrothecae merely comprising pedicels of varied length, each with a proximal bulge, otherwise smooth, arising from lateral wall of a lower pedicel at a point below hydrothecal diaphragm. Hydrothecae fairly shallow, much wider than deep, hydrothecal walls curving outwards from diaphragm to margin, hydrothecal margin strongly flaring, base of hydrotheca with distinct diaphragm below a ring of prominent desmocytes; hydrothecal depth 0.03–0.05 mm, diameter at margin 0.12–0.16 mm, at diaphragm 0.09–0.11 mm. Perisarc of moderate thickness. Hydranths much larger than hydrothecae; hypostome conical; tentacles filiform, about 20–25 in number, in one whorl, armed with small microbasic mastigophores and larger pseudostenoteles. Nematophores, nematothecae, nematodactyls, apical stolons, opercula, pseudodiaphragms, and zooxanthellae absent.
Gonophores fixed sporosacs, encased by solitary gonothecae arising from stolon, from a hydrophore proximal to hydrotheca, or directly from within a hydrotheca; gonothecae solitary, not part of a glomulus. Female gonothecae irregularly obovate, tapering proximally, widest near distal end, both walls convex but side opposite orifice more curved distally; orifice distolateral, accommodating two protruding hydranths; orifice comprising a pair of modified hydrothecae with a ring of desmocytes just below rim; adjoining adthecal process prominent; gonothecae 0.7 mm long, maximum width 0.4 mm, laterally flattened, with no annulations, crease lines, or latitudinal grooves, containing 4– 6 eggs or embryos. Male gonophores not seen.
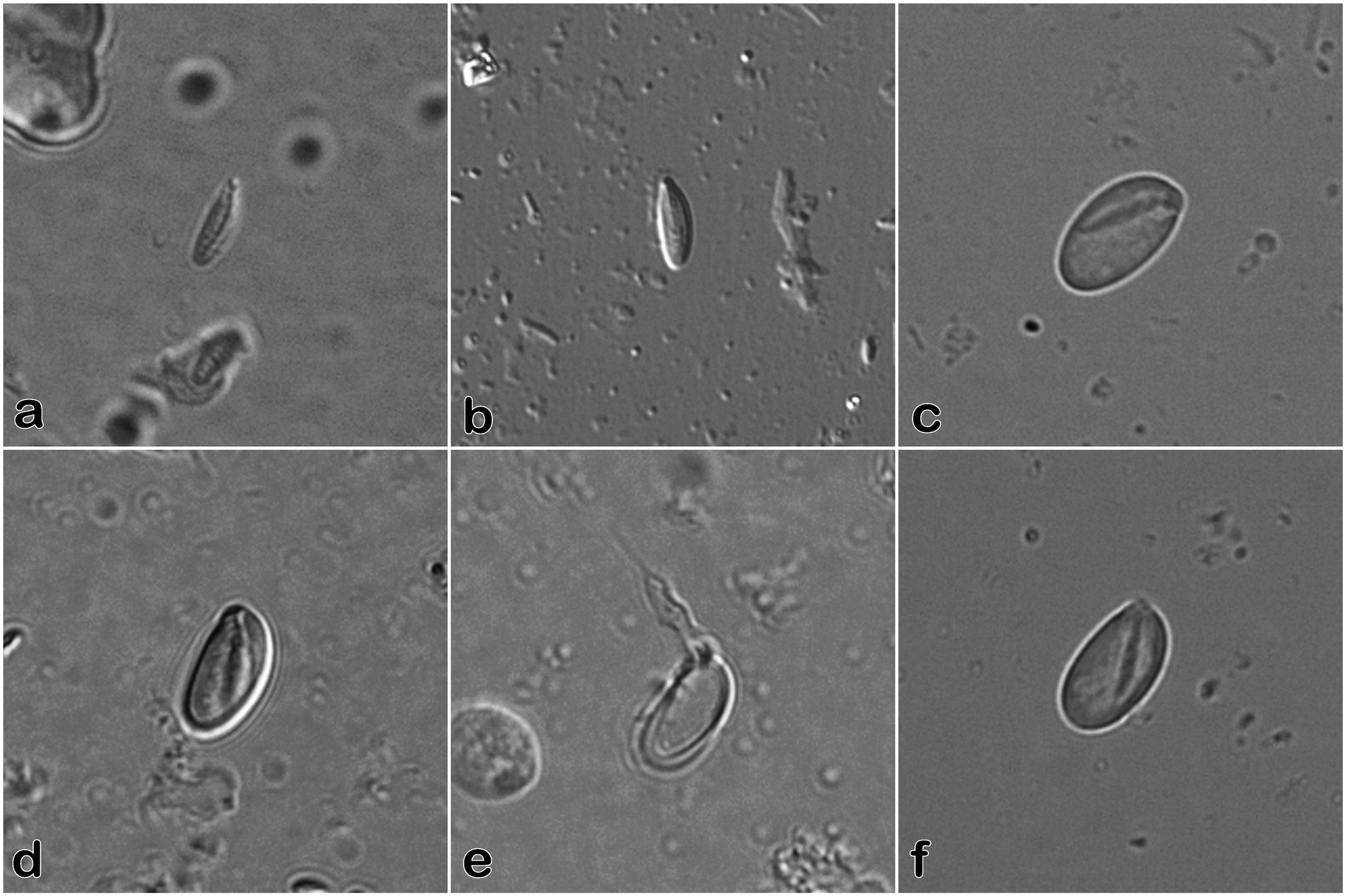
Cnidome ( Fig. 20 View FIGURE 20 ) Hydranths—
microbasic mastigophores (n = 15): 5.9–6.8 µm long × 1.7–2.2 µm wide (undischarged)
pseudostenoteles (n = 13): 9.0–10.0 µm long × 3.9–5.1 µm wide (undischarged)
Hydranths issuing from female gonothecae—
microbasic mastigophores (n = 15): 6.0–6.8 µm long × 1.7–2.4 µm wide (undischarged)
pseudostenoteles (n = 6): 9.1–10.0 µm long × 4.7–5.4 µm wide (undischarged)
Remarks. As with the genus Eudendrium Ehrenberg, 1834 discussed above, synonymies of species of Halecium Oken, 1815 are seldom complex because gonophores are fixed, and no free medusa stage is known to occur in the group. Yet the genus is problematic because of the large number of species assigned to it (>100), the limited number of taxonomically useful characters available for scrutiny (including the lack of a medusa stage), the variations in morphology that occur intraspecifically, and the incompleteness of some early descriptions, especially accounts of putative species based on infertile specimens. Recent attention has been given to the nematocyst complement of species within the group, but the cnidome has been examined in so few species to date that it is presently of limited aid in identification.
A character of prime importance in differentiating species of Halecium is the morphology of the female gonothecae. In H. halecinum ( Linnaeus, 1758) , type species of the genus, and in at least 39 other species besides H. praeparvum ( Table 3), such gonothecae may be broadly described as irregularly obovate, bilaterally symmetrical, and with an aperture for hydranths that is lateral rather than terminal. By contrast, female gonothecae in many other species of the genus are lenticular structures of varied shape and ornamentation that have an aperture at the distal end. Whether these differences are of taxonomic significance at the generic level remains to be explored in detail and confirmed through molecular studies.
Within the group resembling H. halecinum in the general form of their female gonothecae, variations of particular note in the structure include presence or absence of: (1) a chimney-like tube or tubes of varied length and shape leading to one or more lateral apertures; (2) longitudinal or latitudinal crease lines on the gonothecal walls; (3) an adthecal process flanking the orifice; (4) aggregated gonothecae in the form of a glomulus (e.g., H. groenlandicum Kramp, 1911 ). In addition, their gonothecae may vary interspecifically from paddle-shaped to reniform to clavate to mitten-shaped to nearly spherical (e.g., H. ralphae Watson & Vervoort, 2001 ), and the location of the aperture may occur laterally, distolaterally, or even proximolaterally (again, see H. ralphae ). Moreover, gonothecal walls may differ in thickness from one side to the other, ridges or crease lines, if present, may vary in extent and prominence, and the walls may be rounded or even polygonal in cross-section. The aperture itself varies from a single or a pair of openings (variably modified hydrothecae) that may be separate or fused, and if fused, they may or may not be separated by a median septum. A hood-like cover over the aperture exists in some species ( H. scutum S.F. Clark, 1877 ), and a wide opening may appear during development along a lateral wall (e.g., H. bermudens e Congdon, 1907). The number of eggs occurring within the gonothecae may also differ from species to species.
Female gonothecae of H. praeparvum ( Figs. 18 View FIGURE 18 a, 19a, b) correspond in morphology with those occurring in the group containing H. halecinum and 39 others ( Table 3). Within this cluster of species, however, only eight [ H. pusillum ( Sars, 1857) ; H. dichotomum Allman, 1888 ; H. mirabile Schydlowsky, 1902 ; H. repens Jäderholm, 1907 ; H. pygmaeum Fraser, 1911 ; H. minor Fraser, 1935 ; H. vasiforme Fraser, 1935 ; H. tabulatum Watson, 2005 ; H. xanthellatum Galea, 2013 ] approach H. praeparvum in having the following combination of characters: (1) hydroids minute; (2) colonies stolonal or if erect, consisting of a short, irregular, monosiphonic series of pedicels and/or hydrophores rather than a continuous hydrocaulus; (3) hydrothecae widening distinctly from diaphragm to margin, and with rims that may be everted. Others in the cluster are unlike the new species described here in one or more of these characters and are excluded from further discussion. Meanwhile, each of the eight similar species listed above differs from H. praeparvum in various ways. For example, in H. pusillum , both pedicels and female gonothecae are strongly annulated, the aperture of the female gonotheca is lateral, and apical stolons are frequent ( Gravili et al. 2015). Halecium dichotomum stands apart in having long, slender pedicels and internodes, occasional dichotomous branching, hydrothecae that are less flaring, and female gonothecae that are prominently annulated, more slender, and possess a lateral aperture ( Allman 1888; Millard 1975). In H. pygmaeum Fraser, 1911 , the orifice of the female gonotheca is a half-moon-shaped opening on the gonothecal wall rather than at the end of a chimney-like tube ( Fraser 1937). Halecium minor Fraser, 1935 (not H. minor Pictet, 1893 ) from Japan is usually stolonal, although erect colonies comprising as many as five pedicels may occur. Its female gonothecae are reniform in shape and resemble those of H. beanii ( Johnston, 1838) in having a lateral aperture ( Fraser 1935). The little-known H. vasiforme , also from Japan, is a species having stolonal colonies, female gonothecae with a lateral aperture, and hydranths with about 16–18 tentacles ( Fraser 1935; Hirohito 1995). Halecium tabulatum from Australia forms small colonies that are sometimes dichotomously branched and lightly polysiphonic basally; the aperture of its female gonotheca is lateral ( Watson 2005). The Caribbean species H. xanthellatum is distinct in having wrinkles at the bases of the pedicels, zooxanthellae in the coenosarc, and female gonothecae that are slender, tubular, and coarsely annulated on the dorsal wall ( Galea 2013). Halecium mirabile from the White Sea and H. repens from the nearby Kola Peninsula have long been regarded as conspecific or likely conspecific ( Linko 1911; Naumov 1960; Antsulevich 2015). As originally described by Schydlowsky (1902), H. mirabile was based on sterile, stolonal material, and its identity is therefore somewhat uncertain. Illustrations of female gonothecae attributed to it in the literature are those of Jäderholm (1909) for H. repens . They are described as being irregularly oval, with about 5–8 semicircular grooves on the dorsal wall ( Naumov 1960; Antsulevich 2015), and there is no prominent adjoining adthecal process as in H. praeparvum . In turn, H. mirabile and H. repens have often been considered synonyms or questionable synonyms of H. curvicaule Lorenz, 1886 ( Dons 1912; Broch 1918; Schuchert 2001a), a hydroid of Arctic and higher boreal waters. That species differs from H. praeparvum in having female gonothecae with a lateral aperture and with pronounced ribs on the dorsal wall, internodes that are long and slender, and hydrothecae with nearly straight walls ( Naumov 1960; Calder 1970; Schuchert 2001b; Antsulevich 2015). Curiously, illustrations of the female gonothecae of hydroids identified as H. curvicaule by Dons (1912: figs. D, G) are much more like those of H. praeparvum than H. curvicaule , although their trophosomes (figs. D–F) resemble the latter in that hydrothecal pedicels are long and hydrothecal walls flare relatively little. Variations attributed by Dons to H. curvicaule seem extraordinarily large for a single species.
Halecium praeparvum View in CoL is clearly distinct from all 59 of the species having female gonothecae of a decidedly different shape than H. halecinum View in CoL ( Table 3), and no further comparisons with them are made here. As for the remaining 40 species included in the genus, female gonothecae are either unknown at present, or the gender of described gonothecae has not been confirmed as female from existing descriptions. However, other morphological characters can be used to distinguish H. praeparvum View in CoL from each of them. From information summarized in Table 3, only eight [ H. humile Pictet, 1893 View in CoL ; H. ornatum Nutting, 1901b View in CoL ; H. diminutivum Fraser, 1940 View in CoL ; H. fragile Hodgson, 1950 View in CoL ; H. perexiguum Hirohito, 1995 View in CoL ; H. pearsonense Watson, 1997 ; H. elegantulum Watson, 2008 View in CoL ; H. tubatum Watson, 2008 View in CoL ] resemble H. praeparvum View in CoL in having minute colonies that are stolonal or if erect, lack a continuous, true hydrocaulus. They also have hydrothecae that are at least somewhat flaring, and hydrothecal rims that may be recurved. Some differences nevertheless exist that differentiate them from H. praeparvum View in CoL . A morphologically simple species, there is little to set H. humile View in CoL apart beyond its tropical distribution, although its hydrothecae are wider at both the diaphragm (0.15 mm) and the rim (0.22 mm) ( Pictet 1893; Schuchert 2003). Halecium ornatum View in CoL is similar in colony form to H. praeparvum View in CoL , but its pedicels are decidedly longer with a few irregularly located nodes, and the rim of the hydrotheca appears to be much more strongly recurved (Nutting, 1 901b). Colonies of H. diminutivum View in CoL are stolonal or minimally erect, with one or two additional pedicels given off in sequence. Notably, all pedicels are long and distinctively annulated or segmented throughout ( Fraser 1940). Halecium fragile View in CoL , also with both stolonal and sparingly erect colonies, differs in having very long, slender, and flexuous pedicels that are marked at irregular intervals by nodes ( Hodgson 1950; Watson & Vervoort 2001). Halecium perexiguum View in CoL is a mostly stolonal species with few characters to differentiate it. However, it appears to differ in having two annulations at the base of its hydrothecal pedicels, renovation of hydrothecae appears to be more extensive, and the appearance of secondary pedicels from the side of the primary ones is very rare (Hirohito 1995). Hydroids of H. pearsonense occur as short (1–2 mm), erect colonies having from 2–7 hydrothecae. The margins of its hydrothecae are not everted, and secondary hydrothecae are infrequent. Internodes and pedicels of the species are often marked by deep ridges ( Watson 1997). Halecium elegantulum View in CoL is a striking species with colonies that are either stolonal, or erect as a linear series of hydrophores each arising from the base of the hydrotheca below. Pedicels and hydrophores are long, very slender, and lack nodes. Hydrothecae are relatively deep and trumpet-shaped ( Watson 2008; Peña Cantero 2014). Colonies of H. tubatum View in CoL resemble those of H. elegantulum View in CoL in having long, slender, cylindrical pedicels, although pedicels may be wrinkled or annulated basally. Its hydrothecae flare strongly from diaphragm to margin, the hydrothecal rim is distinctly everted, and secondary hydrophores and hydrothecae occasionally arise from within the hydrothecal cavity ( Watson 2008; Peña Cantero 2014). Finally, one other species with undescribed female gonothecae, Halecium exaggeratum Peña Cantero, Boero & Piraino, 2013 View in CoL , deserves mention. While its colonies are also small (up to 15 mm high), a continuous hydrocaulus with long, thin internodes readily distinguishes it from H. praeparvum View in CoL .
genus, H. halecinum View in CoL (i.e., irregularly obovate, bilaterally symmetrical, presence of a lateral opening for one or more hydrothecae); “dissimilar” = gonotheca distinctly different from that of halecinum View in CoL ; “unknown” = female gonotheca unknown or identity uncertain. Under “Colony Form,” “CT” = colony tiny (<1 cm), “CS” = colony small (1–5 cm), “CL” = colony large (>5); “M” = hydrocaulus typically monosiphonic or without a true hydrocaulus, “P” = hydrocaulus typically polysiphonic, “S” = colony stolonal; “IL” = stem internodes long, “IM” = stem
internodes medium length, “IS” = stem internodes short, “IV” = stem internodes highly varied. Under hydrothecal shape, “RR” = hydrotheca strongly flaring with margin often recurved, RS” = hydrotheca not strongly flaring and margin infrequently recurved; “HD” = hydrotheca relatively deep, “HS” = hydrotheca shallow. Halecium flexile Allman, 1888 , not listed in
WoRMS, was recognized as valid following Galea (2010a) and Galea et al. (2014). Also included herein is Halecium pygmaeum Fraser, 1911 because of its morphological similarity.
……continued on the next page TABLE ³. (Continued) ……continued on the next page TABLE ³. (Continued) ……continued on the next page TABLE ³. (Continued) ……continued on the next page TABLE ³. (Continued) Weismann (1883) established this name as “ Halecium tenellum, var. meditteranea .” The variety was elevated to the rank of species by Stechow (1919), as H. mediterraneum , not
meditteraneum as per the original spelling. Prevailing usage, following Stechow and others, has been maintained here.
The name Halecium minor Fraser, 1935 is an invalid junior homonym of H. minor Pictet, 1893 . A replacement name, H. permodicum , has been proposed for the junior homonym herein.
Nutting (1905: 942) considered it possible that a medusa might be produced in Halecium scandens . Also, the lenticular gonophores arose from the hydrothecal cavity rather than from the
stems or hydrorhiza, as in Sagamihydra Hirohito, 1995 . The generic affinities of H. scandens remain uncertain.
The genus Halecium is well-represented in the Bay of Fundy, with 13 species besides H. praeparvum having been reported from the area (Appendix 1). Most common of these are the typically boreal hydroids H. articulosum S.F. Clark, 1875 , H. beanii ( Johnston, 1838) , H. halecinum ( Linnaeus, 1758) , and H. muricatum ( Ellis & Solander, 1786) . Their presence reflects the boreal character of the bay and its waters.
Halecium praeparvum was found during the summer of 2005 on Laminaria growing just below the surface of the water on a salmon aquaculture pen near Eastport, ME. It was inconspicuous because of its diminutive size, and obscure in growing amongst tangles of the algal holdfast. Salinities in the area are thought to have been in the euhaline range (>30‰).
Finally, a nomenclatural problem exists with the binomen Halecium minor ( Table 3). With H. halecinum var. minor Pictet, 1893 justifiably elevated to the rank of species, as H. minor Pictet, 1893 , the name Halecium minor Fraser, 1935 becomes an invalid junior homonym of it. A replacement name for the junior homonym, H. permodicum , nom. nov., is proposed here. The specific name is derived from the Latin adjective permodicum , meaning “very small”.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Hydroidolina |
|
Order |
|
|
Family |
|
|
Genus |
Halecium praeparvum
| Calder, Dale R. 2017 |
Halecium exaggeratum Peña Cantero, Boero & Piraino, 2013
| Pena Cantero, Boero & Piraino 2013 |
H. elegantulum
| Watson 2008 |
H. tubatum
| Watson 2008 |
H. pearsonense
| Watson 1997 |
H. perexiguum
| Hirohito 1995 |
H. fragile
| Hodgson 1950 |
H. diminutivum
| Fraser 1940 |
H. ornatum
| Nutting 1901 |
H. humile
| Pictet 1893 |
Halecium
| Oken 1815 |