
Antillopsyche sessilis, Aguila, Rayner Núñez & Davis, Don R., 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4066.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:18A89A9A-A00C-46E4-BD4C-93084DE5B957 |
|
DOI |
https://doi.org/10.5281/zenodo.6062525 |
|
persistent identifier |
https://treatment.plazi.org/id/03A5294F-FFA5-FFF5-26C1-C730FAE16A23 |
|
treatment provided by |
Plazi |
|
scientific name |
Antillopsyche sessilis |
| status |
sp. nov. |
Antillopsyche sessilis sp. nov.
Male ( Figure 8 View FIGURES 8 – 9 ). Forewing length: 5.2–6.6 mm.
Head: Vestiture white. Antenna 0.3–0.4x length of forewing, 36–43 segmented; scales white; remainder of flagellum brown. Labial palpus entirely white, some individuals with exterior of basal segment brown and scattered scales of the same hue at second segment.
Thorax: White. Forewing background shiny cream, with suffusion of pale ochreous scales more concentrated at base; a row of ochreous spots along fold, at apex, and in a transverse band from two-fifths of costa to termen; small darker spots at termen and at basal one fifth of fold; fringe shiny whitish cream. Venter with same pattern but indistinct, basal one fourth of costa brown. Hindwing shiny whitish cream, semitransparent. Legs white, fore and hind tibia hairy, middle tibia with a lateral brush.
Abdomen: Vestiture shiny, whitish cream.
Genitalia: As described for genus.
Female ( Figure 9 View FIGURES 8 – 9 ). Forewing length: 8.7 mm. Similar to male except: antenna 0.25x length of forewing. Forewing (partially rubbed) with darker spots extending to base, the dorsal two- thirds of transverse band and the basal half of fold. Frenulum consisting in a cluster of 5 smaller bristles.
Abdomen: shiny whitish cream, scaling more dense laterally; corethrogyne scales slightly darker.
Genitalia: as described for genus.
Larva ( Figures 10–20 View FIGURES 10 – 20 ). Maximum length examined 10.2 mm; maximum head width ~ 1.2 mm. Body cuticle dark brown with pinnacula much paler in color. Head: Epicranial notch moderately shallow, ~ half the length of epicranial suture ( Figure 14 View FIGURES 10 – 20 ). P1 arising closer to AF2 than A1. AF2 slightly longer than AF1 ( Figure 14 View FIGURES 10 – 20 ). Five stemmata present and evenly spaced in an anteriodorsal arch; stemmata 4–5 immediately below 3 ( Figure 14 View FIGURES 10 – 20 ). Antenna and maxilla with sensilla as shown ( Figures 15 & 17 View FIGURES 10 – 20 ). Labrum with six pairs of dorsal setae, all of moderate length except for elongated M3 and La2; venter of labrum with three pairs of epipharyngeal setae, the two mesal setae only slightly longer than lateral seta ( Figures 18–19 View FIGURES 10 – 20 ). Mandible with five cusps which become smaller mesally; lateral margin serrated basal to lateral cusp ( Figure 20 View FIGURES 10 – 20 ). Labial palpus with basal segment (1) very elongate, ~ 2/3 the length of spinneret; segment 2 greatly reduced, ~ 0.1 the length of segment 1; apical seta from segment 2 long, ~ 2x the length of segment 2. Spinneret long and slender, gradually tapering to narrow, simple apex. Thorax: Pronotum completely fused with prespiracular pinaculum bearing L1–3 and spiracle; XD1 moderately long, ~ 1.2x the length of XD2 ( Figure 13 View FIGURES 10 – 20 ); subventral pinnaculum well developed, extending nearly the length of segment. Dorsal pinnacula of mesothorax fused to form single, light brown plate nearly as large as pronotum; dorsal pinnaculum of metathorax also connected mid-dorsally but divided laterally with SD1 and 2 on lower pinnaculum; lateral pinnacula narrowly separated from notal plates. SV bisetose on T1–3 ( Figure 13 View FIGURES 10 – 20 ). Abdomen: D1 and D2 arising from separate oval pinnacula on A1-9 ( Figures 12–13 View FIGURES 10 – 20 ); pinnacula bearing D1 on A1 and A2 fused with opposite pinnacula dorsally; pinnaculum of D2 indistinct on A9 ( Figures 13 & 16 View FIGURES 10 – 20 ); SD bisetose on A1-8 with SD2 minute, SD2 absent on A9; L series trisetose on A1–9 with L3 arising from separate pinnacula on A1–7, and L1–3 each from separate pinnacula on A8–9 ( Figures 13 & 16 View FIGURES 10 – 20 ); SV bisetose on A1-2, 7–9, trisetose on A3–6 ( Figure 13 View FIGURES 10 – 20 ). Anal plate with seta D2 absent, represented instead by prominent pore as in Brachygyna (Davis 1999) ( Figure 16 View FIGURES 10 – 20 ); prolegs 3–6 with 20–26 crochets in a uniordinnal, lateral penelipse. Anal crochets ~ 21 in a uniordinal row opened caudally. Cuticle surrounding anal aperture thickened but unmodified and similar in texture to remainder of segment.
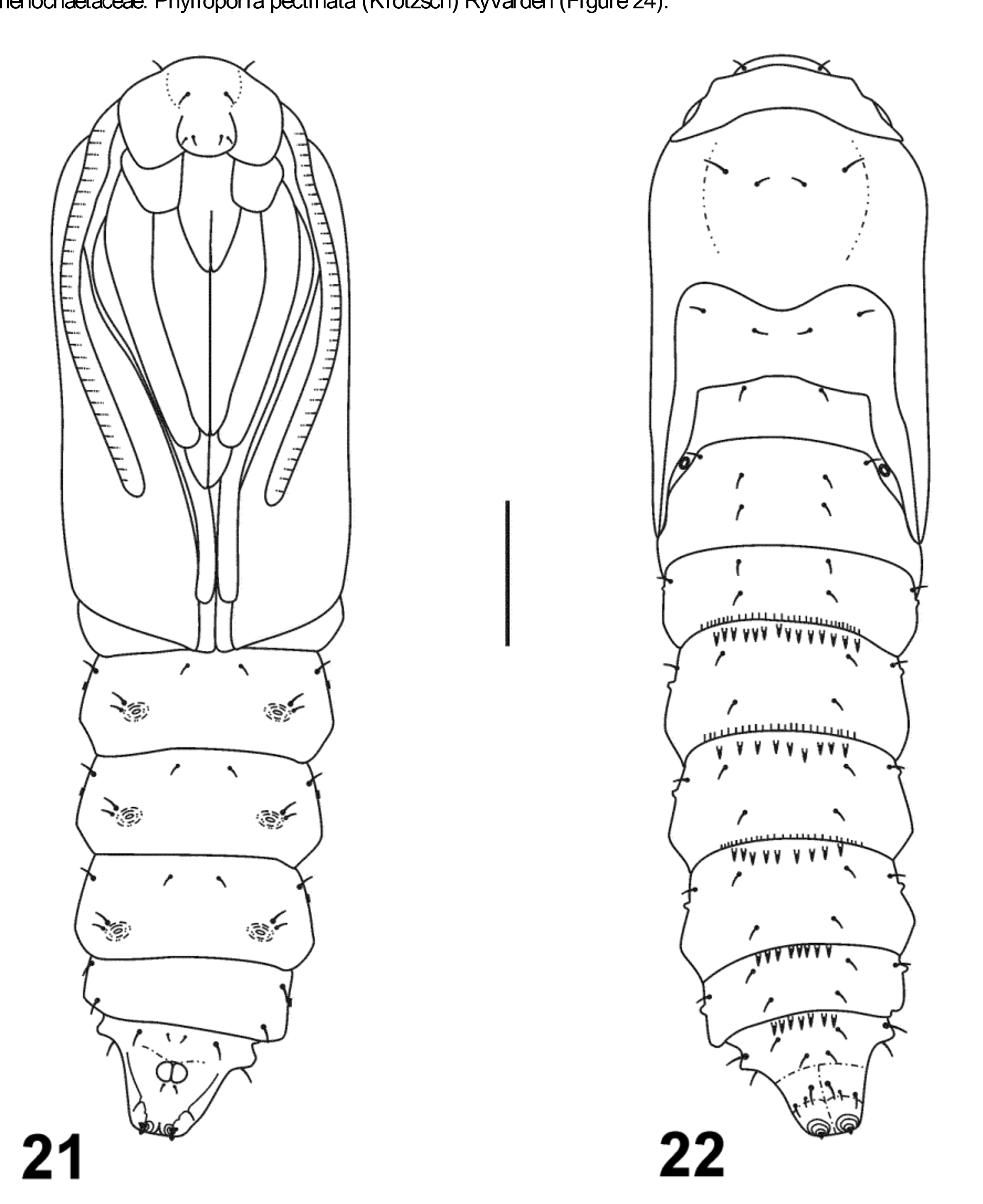
Male pupa ( Figures 21–22 View FIGURES 21 – 22 ). Length 5.3–7.0 mm, maximum width 1.3–1.9 mm (n=7). Color amber in smaller individuals, pale reddish brown in larger ones. Wing sheaths reaching anterior margin of abdominal segment A4 ( Figure 21 View FIGURES 21 – 22 ). Spiracle of A1 exposed close to hindwing margin. Head with one pair of dorsal setae, one pair on vertex and two pairs on frons; two pairs of dorsal setae on pronotum, mesonotum and abdominal segments A1–A8 ( Figure 21 View FIGURES 21 – 22 ); A8 with three additional pairs of setae anterior to cremasteral spines. Venter with SV series bisetose on A4–A6, setae close together and to proleg scars ( Figure 22 View FIGURES 21 – 22 ); A8 with four pairs of setae: two near anterior margin, one posterior to protuberances and one on ridge edges of ventral pair of cremasteral spines ( Figure 22 View FIGURES 21 – 22 ). Dorsum of abdomen with a single row of posterior oriented spines on anterior margin of A4–A8 and a single row of anterior oriented spines on posterior margin of A3–A5 ( Figure 22 View FIGURES 21 – 22 , Table 1). Cremaster consisting of a pair of adjacent round tubercles at middle of segment and two pairs of small spines, one pair directed ventrally and the other, at apex, directed caudad; anal groove indistinct.
Female pupa. Length 8.5–9.4, maximum width 2.2–2.4 (n=2). Similar to male except: abdomen enlarged; average number of spines on dorsum of abdomen larger, spines absent from anterior margin of A8 or represented by a single large spine with a broad base (Table 1).
Larval case ( Figures 23–24 View FIGURES 23 – 24 ). Brown, cylindrical; largest case 31 mm in length, 4 mm in diameter; walls soft, covered externally with fecal pellets and minute fragments of bark and fungi, lined internally with dense grayish white silk.
Type material. Holotype: ♂, CUBA: Granma, Bartolomé Masó, La Aguada de Joaquín, 1300 m, 2.xii.2007, ex Corticium sp., emerged 24.i.2008, (R. Núñez) ( CZACC). Paratypes: 6 ♂, 1 ♀. 3 ♂, same data as holotype except: emerged 23.i.2008, 24.i.2008, and 8.ii.2008, ex Corticium sp., ex Phylloporia pectinata , and Trametes villosa . ♀, same data as holotype except: 12.ii.2007, ex Corticium sp., ex Phylloporia pectinata , and Trametes villosa ; emerged 25.iii.2007, (R. Núñez & E. Fonseca). 3 ♂, same data as anterior except: emerged 24.iii.2007, emerged 4.iv.2007, emerged 5.iv.2007. 3 larval cases with associated pupal exuviae or pupal remains, CUBA: Granma, Bartolomé Masó, camino al Pico Turquino, alrededores de La Platica, 1000 m, 22.vii.2009, (R. Núñez) ( CZACC, USNM).
Hosts ( Figures 23–24 View FIGURES 23 – 24 ). Corticiaceae : Corticium sp.; Polyporaceae : Trametes villosa Fr. (Kreisel) ( Figure 23 View FIGURES 23 – 24 ); Hymenochaetaceae : Phylloporia pectinata (Klotzsch) Ryvarden ( Figure 24 View FIGURES 23 – 24 ).
Distribution ( Figure 25 View FIGURE 25 ). Known only from three adjacent montane localities in the Sierra Maestra of southeastern Cuba.
Etymology. The specific name is derived from the Latin sessilis (sitting) in reference to the sedentary feeding behavior by the larva of this species, unusual among known Psychidae .
Habitat. Antillopsyche sessilis inhabits the rainforests and evergreen forests at intermediate elevations, 1000– 1300 m. The relative humidity of this region is between 87 and 92% in the morning (7:00 am) and between 75 and 80% in the afternoon (1:00 pm) ( Montenegro 1991). The precipitation is between 1,800 and 2,300 mm per year. Average annual temperature varies between 16 and 20ºC. January average temperatures fluctuate between 14 and 18ºC, and July temperatures between 18 and 22ºC.
Flight period. Reared specimens emerged from January to April. Because data are from only two collections (February and December) there is little to report on seasonal occurrence. However, high humidity and relative low temperatures of habitat through the year probably provide suitable conditions for development in every month.
Biological observations. The species feeds on a wide variety of fungal hosts. On one standing dead tree at La Aguada de Joaquín, several larvae were found feeding on the three hosts mentioned above. The larval feeding behavior of A. sessilis differs from that commonly found throughout the Psychidae in being more restricted and attached to a specific site. They probably also feed inside the fungi but it seems to occur only during the initial stages. Even larvae with cases directly attached to fungi were observed eating on the external surface ( Figure 24 View FIGURES 23 – 24 ). This may be a strategy to obtain food when it is scarce by increasing the foraging area for each individual, as shown Fig. (23) where one larva is feeding on a trunk with all Trametes villosa almost completely devoured. The larva tunnels into fungus and/or wood and firmly attaches the case to the entrance. The interior of the case and the tunnel substrate are covered with the same continuous silk layer. All cases with living individuals were found on standing dead trunks ( Figure 26 View FIGURES 26 – 27 ). A small group of old empty cases was collected on a fallen dead trunk a few inches above the ground. The distance from the ground of all observed or collected specimens varied between 0.5 to 2.5 meters.
Larvae were observed feeding both during the day and night in nature and in the laboratory. Larvae were not observed feeding at the proximal (anterior) end of case as do nearly all other Psychidae . Instead, they are in an inverted position and foraging at the distal end of the case onto the external surface of the tree trunk (left arrow in Figure 26 View FIGURES 26 – 27 ). When disturbed, the larvae retreat rapidly to the safety of the tunnel within the trunk.
Before pupation, the larva closes the distal end with silk forming a slit-like opening by flattening the transverse section of the case. Although the exact site of pupation was not observed, it seems to occur inside the tunnel or outside close the entrance. The reason for this uncertainty is that two larval cases were bent at the distal two-fifths after pupation and both adults emerge from the angle (left case at Figure 27 View FIGURES 26 – 27 ). Thus, pupation probably occurs at least in one of the two locations suggested above. The bending of the case may have been caused by a species of Dryadaula (Dryadaulidae) ( Figure 28 View FIGURES 28 – 29. 28 ). Only a single female of Dryadaula emerged few days prior to the emergence of the A. sessilis adult, from one of the two bent cases.
The pupal stage requires 32 to 42 days (n=3) and emergence occurs shortly after sunset (8:00 to 9:00 pm, n=3). Although night collecting with mercury vapor lights was conducted at both localities, no adults were observed or collected. A 250-watt bulb on a sheet was used during 12 nights between January 2005, February 2007 and November 2007 at La Aguada de Joaquín. This method was followed at La Platica, a small village just 300 m away from the light collecting locality, during 20 nights ( January 2005, February 2007, November 2007, August 2008, May 2009 and July 2009) without the sighting or collecting of a single specimen.
Larvae collected at La Aguada de Joaquín were attacked by parasitoids. Several wasps emerged from two larvae were identified as a species of Baryscapus Förster, 1856 ( Eulophidae : Tetrastichinae ) ( Figure 29 View FIGURES 28 – 29. 28 ).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Tineoidea |
|
Family |
|
|
Genus |