
Henricia pumila, Eernisse, Douglas J., Strathmann, Megumi F. & Strathmann, Richard R., 2010
|
publication ID |
https://doi.org/10.5281/zenodo.275445 |
|
DOI |
https://doi.org/10.5281/zenodo.6205509 |
|
persistent identifier |
https://treatment.plazi.org/id/03A0E238-AE7D-FFFD-FF42-07FBFDB32F3D |
|
treatment provided by |
Plazi |
|
scientific name |
Henricia pumila |
| status |
sp. nov. |
Henricia pumila View in CoL , n. sp.
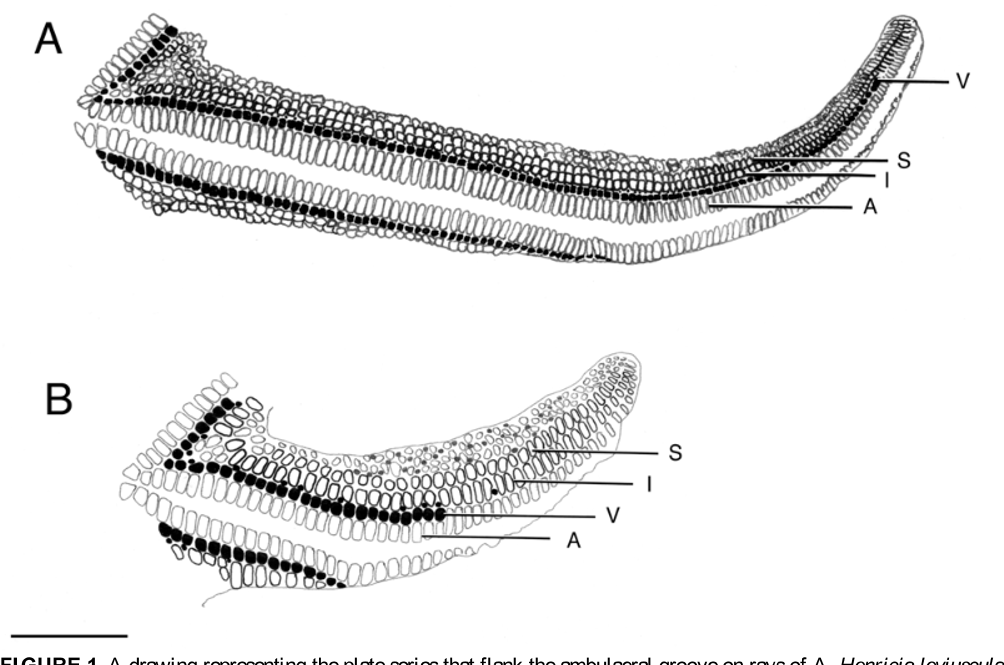
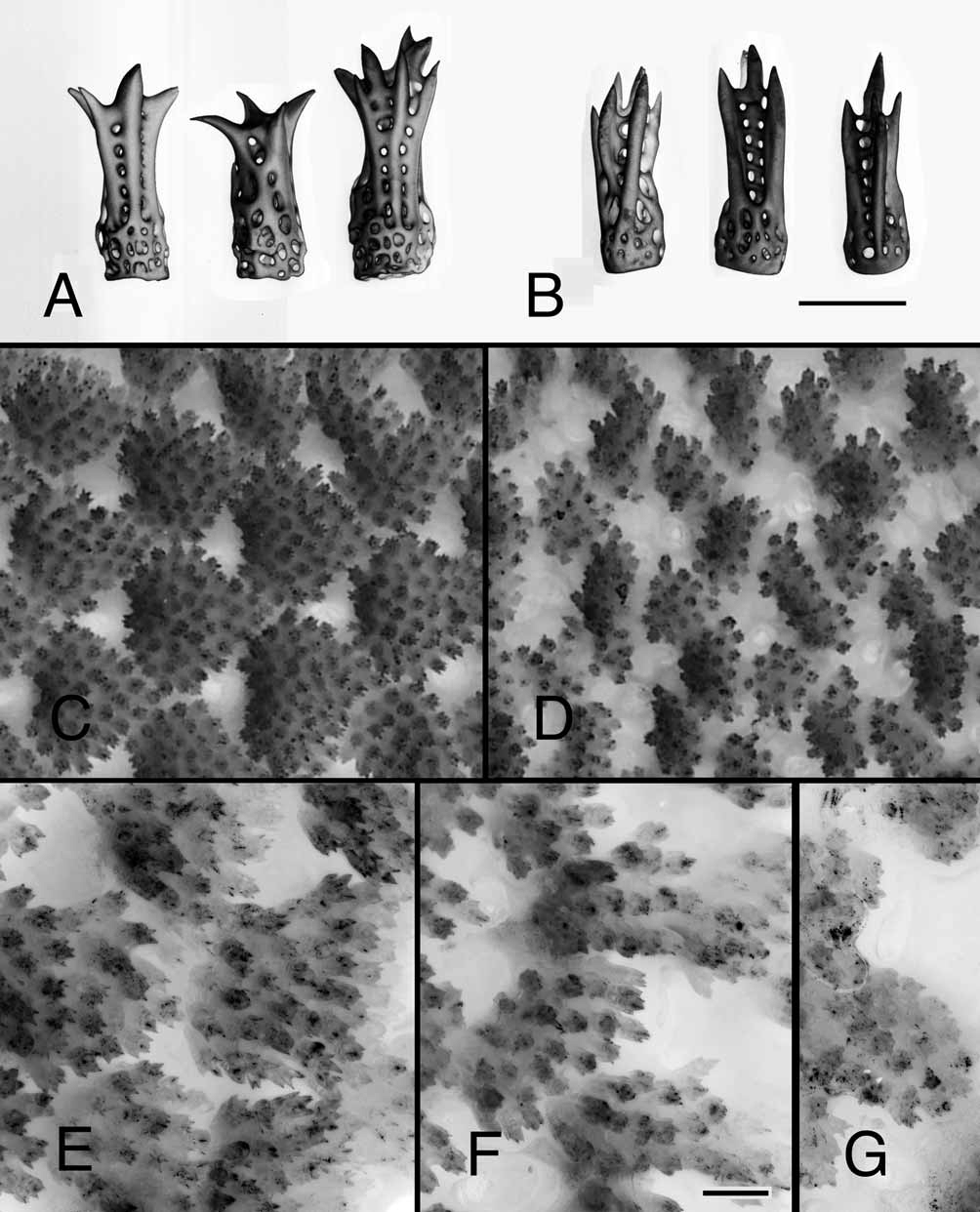
Figs. 1 View FIGURE 1. A B, 3B–D, 4B–C, 5B, 5D, 5F–G
Type Material: Holotype USNM 1116585; paratypes USNM 1116586; CASIZ 180559 and 180560; LACM 1987- 519.001 and 2005-065.001; MFS 109
Type Locality: Mar Vista Resort, San Juan Island, Washington, USA (48°28.595ʹN 123°04.015ʹW), rocky intertidal under rock in tidepool
? Cribrella laeviuscula View in CoL var. crassa H.L. Clark 1901:328
Henricia leviuscula View in CoL variety F, in part, Fisher 1911 (see also Feder, 1980; Lambert, 1981; Mah, 2007) Henricia leviuscula View in CoL ( non Stimpson, 1857); in part, Hopkins, 1967: 19 –23, 65–68
Other material examined: More than 100 specimens in our collections from the San Juan Archipelago, San Juan Co., Washington, plus other specimens from southern Vancouver Island, British Columbia, Canada, intertidal; Bodega Head, Sonoma Co., California, intertidal; Franklin Pt., San Mateo Co., California, intertidal; Monterey Breakwater, Monterey Co., California, approx. 12–15 m, including one brooding female; Shelter Cove, between Shell Beach and Pismo Beach, San Luis Obispo Co., California, intertidal; Arbolitos, south of Punta Banda, Baja California, Mexico, intertidal. Museum specimens identified included: USNM 1083883 ( 3 in lot), Carmel, Monterey Co., California; USNM E3831 ( 2 in lot), Monterey Bay, California; USNM E3832 ( 2 in lot), Strait of Juan de Fuca, Washington; USNM E21533, San Nicolas Id., California; USNM 1083885 ( 3 in lot), Puget Sound, Washington; CASIZ 0 0 8558 Crescent City, California ( 1 specimen in alcohol, likely “variety F” voucher for Fisher, 1911:285); LACM 1959-281.12 ( 1 specimen), Monterey Co., California, intertidal, 36°32’N 121°56’W; LACM 1928-3.1, Pacific Grove, California, intertidal. We have additionally examined images of seastars from the vicinity of Sitka, Alaska ( Fig. 3 View FIGURE 3 C) and from Cape Arago, Oregon ( Fig. 3 View FIGURE 3 D) that appear to be the same species.
Diagnosis. Small in size; rays stout and short (R/r less than 5), aboral pseudopaxillae well-spaced, bearing up to 50 short spines with fenestrated, crystalline, smooth-sided shafts tipped by up to 10 heavy sharp points that do not noticeably splay. Aboral color in life usually a mottled pattern of ochre, brown, gray, rust-red, or yellow; oral color yellow to cream. Gonopores orally-directed, eggs non-buoyant, development benthic and brooded.
Description. Holotype ( Figs. 3 View FIGURE 3 B, 4B, 5D, 5F), collected by RRS and MFS on March 30, 2005, is a medium-sized specimen, in life R = 19.7 mm, r = 6.4 mm. Madreporal ray 6.8 mm wide at base. Living aboral colors are ochre mottled with red-brown spots on every ray, orange-red around the anus extending toward one interradius, and orange at the ray tips ( Figs. 3 View FIGURE 3 B). Madreporite cream yellow. Oral color orange yellow. A large aboral pseudopaxilla with 25 spines ( Fig. 4B View FIGURE 4. A – B , 5 View FIGURE 5. A – B D, 5F).
Most specimens have 5 rays; 4- or 6-rayed individuals are rare. The rays taper rather evenly from base to tip but are short and stout. Among 34 specimens from the San Juan Archipelago (the largest having R = 29.4 mm and r = 6.3 mm and the smallest having R = 4.1 mm and r = 1.4 mm), R/r varied from 1.8 to 4.7, average 3.0. Aboral color usually mottled, the mix may include yellow, ochre, brown, rust-red and gray, all colors not always present on each individual (southern specimens show a greater range of mottling); oral color cream to orange yellow. Madreporite distinct; madreporal spines grouped at the periphery and in single rows across the center and the same size as the aboral pseudopaxillar spines. Aboral pseudopaxillae well-spaced and separated by recessed tissue and papular areas containing 1 to 4 papulae, the maximum number increasing with body size. Aboral pseudopaxillae oval or irregularly oblong on the disc and oval to crescentic on the rays consisting of tissue-covered spines in groups of 2 to 50, usually 12 to 45, the number increasing with body size.
Aboral spines are short and stout and terminate in multiple short sharp points that do not markedly splay and so are directed more or less distally ( Figs. 5B, 5 View FIGURE 5. A – B D, 5F), which can be obscured by tissue in living or airdried ( Fig. 5 View FIGURE 5. A – B G) specimens. On the rays, two marginal and one ventrolateral series of pseudopaxillae form regular and obvious rows flanking the ambulacral plates that edge the ambulacral furrow. Near the ray base, a small triangle of interradial pseudopaxillae is enclosed between the superomarginal series, as it descends from the aboral disc to lie along the ray side, and the inferomarginal series. Both the supero- and inferomarginal pseudopaxillae are slightly larger than the aboral pseudopaxillae and extend to the ray tip in 1:1 correspondence with the ambulacral plates. The ventrolateral pseudopaxillae are small and this series extends half to three-fourths of the length of the ray ( Fig. 1 View FIGURE 1. A B, 4C). Among specimens with R ranging from 9.0 to 29.4 mm, the ratio of ventrolateral to adambulacral pseudopaxillae (V/A, counted from mouth to ray tip) ranged from 0.34 to 0.79, averaging 0.62 (N = 53; holotype V/A = 0.74).
A regular series of single papulae occurs between the inferomarginal and ventrolateral pseudopaxillae and single papulae between some ventrolateral and adambulacral pseudopaxillae.
Near the base of the ray, superomarginal pseudopaxillae have 12–37 spines each; inferomarginals, 10–32; and ventrolaterals, 5–20. Number of spines per pseudopaxilla generally increases with body size. Each adambulacral plate bears one thin, curved deep-furrow spine and 6–14, usually no more than 10, large, slightly curved, columnar spines in one row that becomes double then triple farthest from the ambulacral furrow. The large spines nearest the furrow are blunt-ended with finely spinous surfaces, the tips sometimes slightly flattened but not spatulate; those farther from the furrow are smaller and more coarsely spinous with sharp terminal points but not radiating thorns. On the holotype, the largest adambulacral spine is about 500 µm long and the points at the tip span 160–200 µm.
Distribution. H. pumila is the only small, brooding species of this genus presently known in Puget Sound and the San Juan Archipelago, Washington. It seems to be a widespread Pacific Coast shallow-water coastal species. In the north, it occurs in southern British Columbia, Canada and probably ranges further north to Sitka, Alaska, USA. The southern-most record is from areas of cold-water upwelling at Arbolitos, south of Punta Banda, Baja California, Mexico, but the species appears to skip over southern California, with the nextmost southern record near Pismo Beach, California, north of Pt. Conception. Despite the lack of records from southern California, it might be present in the Channel Islands or in unexplored subtidal areas.
Reproduction. We have assumed sexes are separate but have not dissected gonads to thoroughly search for evidence of hermaphroditic tissues, as is known for some small brooding marine invertebrates (e.g., Strathmann et al., 1984; Eernisse, 1988; Colgan et al., 2005; Keever and Hart, 2008). Eggs are shed through gonopores on the oral side of the disc edge between rays ( Chia, 1966; Hopkins, 1967), in contrast to their aboral position in H. leviuscula and other free-spawning species. Eggs about 1144 µm diameter ( Strathmann et al., 2002) are neither buoyant nor sticky when shed and are held beneath the maternal body with the rays spiraled, pinwheel fashion, around a slightly elevated disc (R. Strathmann, unpubl. observ.). Embyos are brooded under the disc and emerge as crawl-away juveniles. Brooding in rocky low intertidal areas of the San Juan Archipelago and central California has been seen in January to April. The Atlantic deep-water congener, Henricia lisa , has recently been shown to be a facultative brooder ( Mercier and Hamel, 2008) and, although the mode of brooding differs, the same possibility exists for H. pumila and other asteroid species known to brood.
Etymology. From the Latin for dwarf, the term used by Fisher (1911) for the small adult body size of this species.
Remarks. It is possible that this species is the same as Hubert Lyman Clark’s (1901) stout-rayed variety Cribrella (= Henricia ) laeviuscula [sic] variety crassa from Puget Sound. Clark was an affiliate of Olivet College, Olivet, MI, about the time he described this variety by only its body size and ray shape. The museum at Olivet College was unfortunately destroyed by fire in 1968, and most of the records were lost (Marie Davis to Robert Woollacott, pers. comm.). We have not found types for Clark’s nominal variety in museums where it seemed plausible that he might have deposited material, including the Museum of Comparative Zoology at Harvard University (where Clark was later curator of echinoderms from 1910 to 1946) or the American Museum of Natural History (Clark reported that the specimens were collected in Puget Sound by a group from Columbia University, and Columbia’s museum later became part of the AMNH collections). A partially dissected dried specimen was found at Harvard’s MCZ; its only label is clearly not the original because it reads “ Henricia laeviuscula crassa (Clark) Locality : Puget Sound, Washington” (MCZ 1046). The morphology of this specimen corresponds to H. pumila but the entries for it in the accession catalogue and a 1930s list of specimens do not give collection information or an accession date. Given the brevity of the description and the lack of verifiable type material, we consider Clark’s varietal name, crassa , to be a nomen dubium, as similarly concluded by Fisher (1911, 1930) and Verrill (1914).
Fisher’s (1911) specimens of the small form that he called H. leviuscula variety F were collected from the vicinities of Monterey, San Francisco, and Crescent City in California and the Straits of [Juan de] Fuca and Puget Sound in Washington. Although Fisher knew some of these to be brooders, and described the brooding posture as arched, his specimens (see also Kozloff, 1996; Mah, 2007) may have included more than the one species we describe here as H. pumila .
There is at least one other at least partly co-occurring, and probably undescribed, small-bodied species from British Columbia south to northern Baja California. Yet another species is usually small and is so far known from the subtidal of central and southern California (D. J. Eernisse and M. Strathmann, in prep.). Both of these species have finer and more numerous spines per plate. Nothing is yet known of their life history traits so the possibility remains that one or both might be brooders. Other Henricia species are known to be brooders but differ in their distribution and morphology. H. tumida Verrill 1909 grows larger and has broader, thicker arms and is reported from the Aleutians Islands, Alaska, and the Bering Sea. It was discussed as H. sanguinolenta eschrichtii or H. tumida by Fisher (1911, 1930), and as H. tumida or H. tumida borealis , or possibly as H. arctica , by Verrill (1909, 1914). Djakonov (1950: 85–86) later described and illustrated specimens of H. tumida , H. tumida borealis , and a similar smaller species, H. arctica , from the Sea of Okhotsk and the Bering Sea. He reported that H. arctica is also found in the Litke Strait and off Cape Lisburne, Alaska. It is not known if H. arctica is a brooding species. Small brooding species have been reported from Japan, and the development of one, H. nipponica , has been described ( Komatsu & Tayayama 1980). We have not studied H. nipponica but believe it unlikely to be the same as H. pumila because nothing resembling H. pumila is yet known from the intervening Aleutian Islands. We have seen little genetic evidence of marine species with such a disjunct eastern and western Pacific distribution. Thus, we speculate that any brooding Henricia in the northwestern Pacific might be closely related to, but not conspecific with, H. pumila . An extension of the study of small brooding species of Henricia to the far northern and northwestern Pacific is still needed.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Henricia pumila
| Eernisse, Douglas J., Strathmann, Megumi F. & Strathmann, Richard R. 2010 |
Henricia leviuscula
| Hopkins 1967: 19 |
Cribrella laeviuscula
| Clark 1901: 328 |