
Empididae, Giebel, 1856
|
publication ID |
https://doi.org/10.11646/zootaxa.1180.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5059456 |
|
persistent identifier |
https://treatment.plazi.org/id/039E87BE-FB10-B256-F11F-F9BD4FC5FA1D |
|
treatment provided by |
Felipe |
|
scientific name |
Empididae |
| status |
|
Empididae View in CoL View at ENA
Empides Latreille, 1809: 189, 191. Type genus Empis Linnaeus, 1758 .
Empididae Giebel, 1856: 206 View in CoL .
The definition of the Empididae View in CoL in this study is more restrictive than in previous classifications. The family is recognized on the basis of the following suite of characters (see also below—Key to Families and unplaced Genus Groups of Empidoidea): Vein R 4+5 normally branched, prosternum enlarged forming precoxal bridge, laterotergite usually with setae, male terminalia often with claspinglike cerci or hypandrium laterally flattened or keellike, female terminalia usually telescopic with simple cerci. No distinct apomorphy is known to define this diverse lineage and further studies on the relationships of all genera in this family are urgently required. This family includes the following generic groupings, tribes and subfamilies:
Incertae sedis within Empididae
Ragas group (includes: Dipsomyia Bezzi , Hormopeza Zetterstedt , Hydropeza Sinclair , Ragas Walker , and Zanclotus Wilder )—This genus group of predaceous empidoids was clearly defined by Sinclair (1999) and is mainly characterized by the recurved labrum and apical epipharyngeal comb. The single tree obtained by a posteriori character weighting using successive approximations, placed this lineage and Brochella in the Empididae as the sister group to the Clinocerinae (Tree 4) primarily on the basis of the presence of a dorsal mesepimeral pocket (character 31.1), reduction of antennomere 10 (character 8.1), and a slender male tergite 8 (character 71.1). However, because of the degree of homoplasy associated with the last two characters in particular, and until the relationships and definition of the entire Empididae are better resolved, the phylogenetic assignment of the Ragas group will continue to remain uncertain.
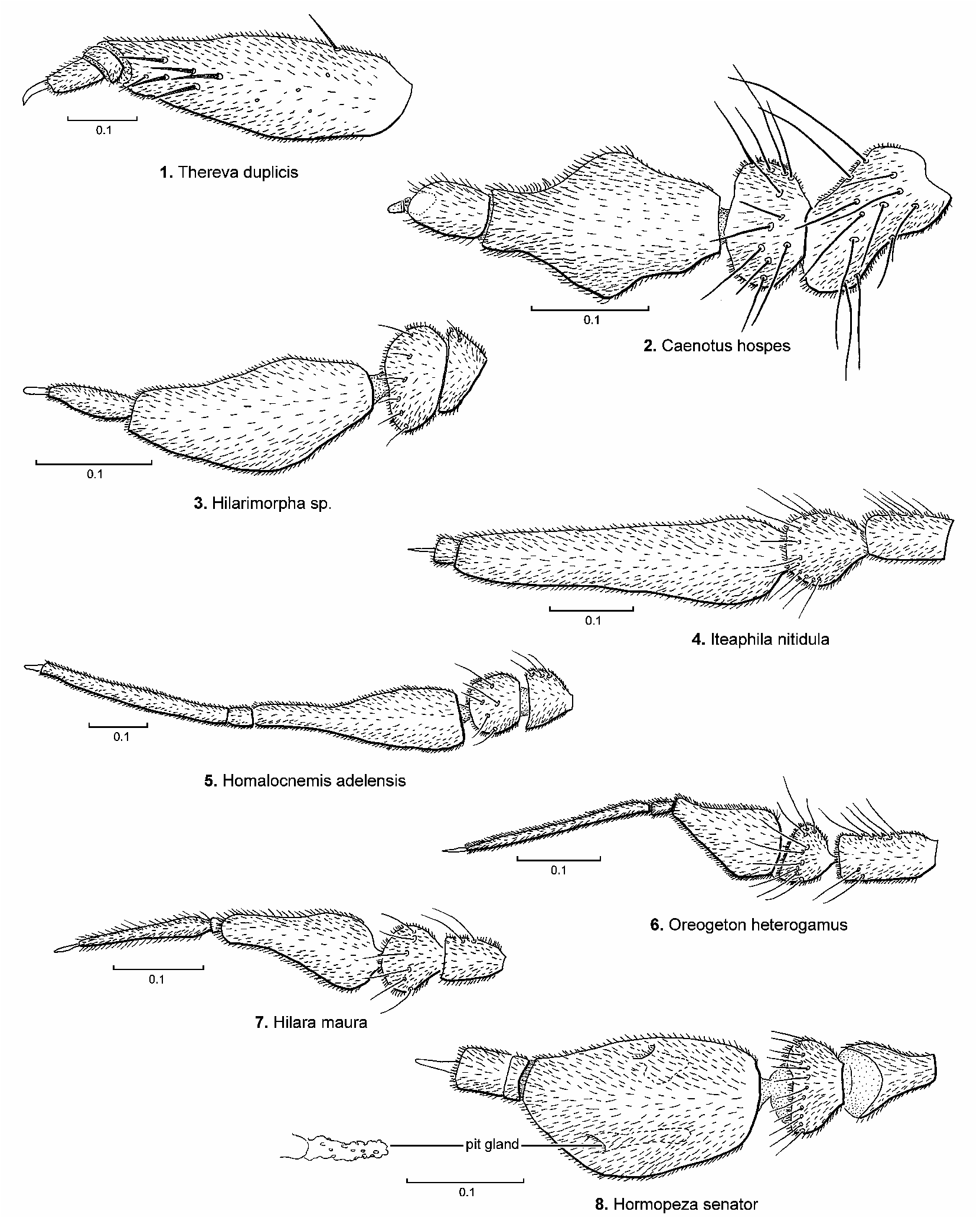
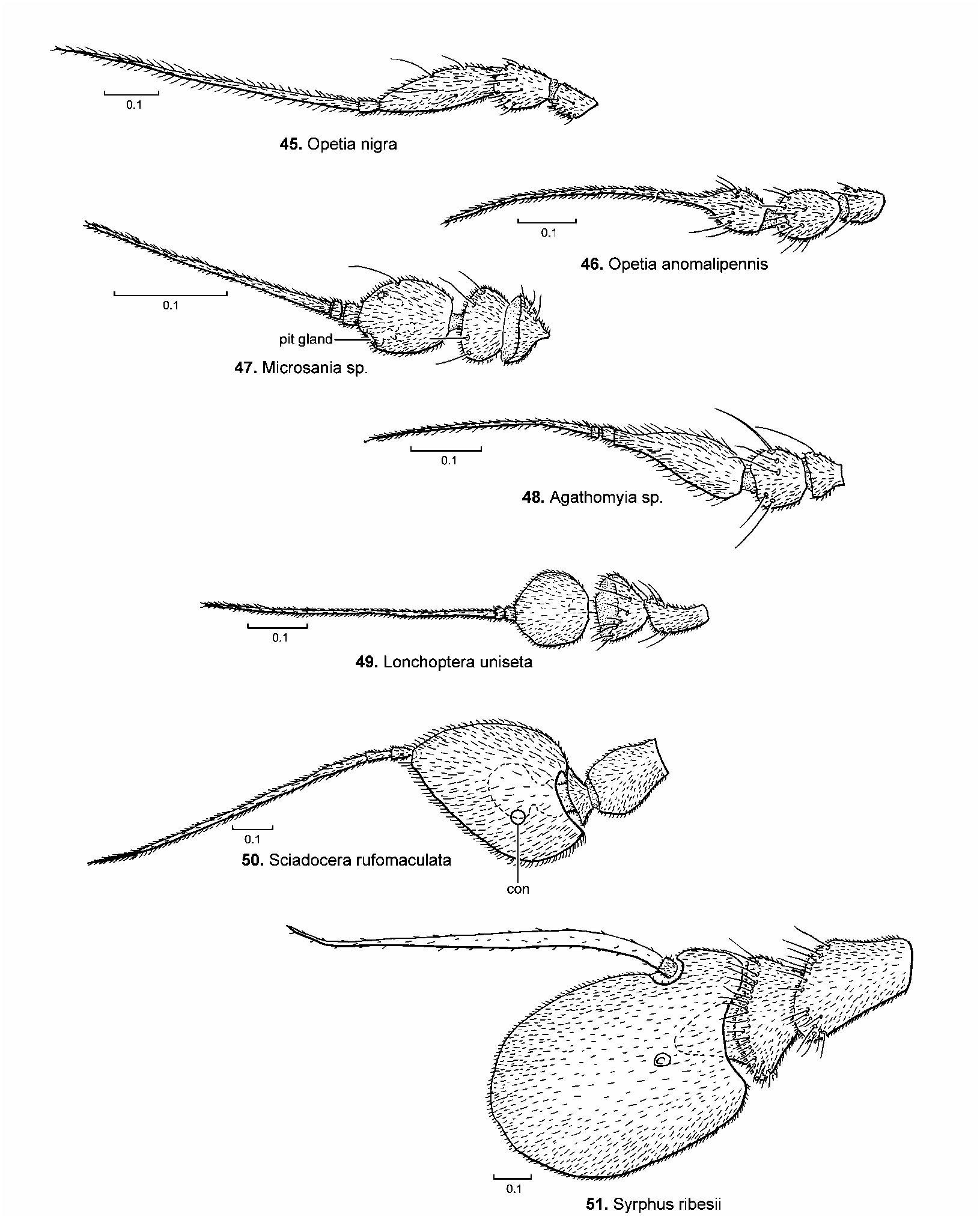
The postpedicel of Hormopeza possesses a pair of sensory pits ( Fig. 8 View FIGURES 1–8 ; Tuomikoski 1960, fig. 1e), which are also very similar in form to those observed in the platypezid genus Microsania ( Fig. 47 View FIGURES 45–51 ). Both genera are commonly known as smoke flies and it is assumed that these pits and inner glands are used in the detection of smoke. Large swarms of both taxa have been observed in smoke from wood fires (see Sinclair 1995b and Bickel 1996b for references). Swarms of Hormopeza usually consist of equal numbers of both sexes, whereas swarms of Microsania consist almost entirely of males.
It has always been assumed that these flies use smoke as an aggregating queue to concentrate both sexes in an area that has been burned over by fires. It is also assumed that certain fungi, which grow only on firescarred wood function as hosts for Microsania larvae ( Kessel 1989: 180). But the immature stages of both taxa remain unknown. It is possible that the smoke from fires does function as an aggregating queue, but mostly to attract the flies to fresh oviposition sites as has been observed for longhorned beetles ( Cerambycidae ), where great numbers swarm about forest fighting crews battling fires in Ontario ( Carney 1999). It is possible that Hormopeza and Microsania also oviposit into the freshly damaged trees. This is a breeding habitat that should be more fully explored.
Brochella Melander —This monotypic genus is an odd unique taxon that occurs in the Cascade Mountains of the western Nearctic Region. Brochella is possibly a flower visitor on the basis of its mouthpart morphology. The single tree obtained by a posteriori character weighting using successive approximations, placed this genus with the Ragas group in the Empididae , as the sister group to the Clinocerinae (Tree 4). However, the phylogenetic assignment of Brochella will remain uncertain, until the relationships of the Empididae are better resolved.
Philetus Melander —This genus includes two species confined to western North America . Very little is known of this genus and most specimens have been sporadically collected. Recently, one species was collected in large numbers during a canopy study of wet temperate rainforests on Vancouver Island (BC, Canada; specimens in CNC).
Hesperempis group (includes: Dryodromia Rondani , Hesperempis Melander , and Toreus Melander )—This genus group represents another flowerfeeding lineage and possibly also includes an undescribed Australian genus. The thoracic chaetotaxy is reduced in all genera, and the bristles are pale white to yellowish.
Schiner (1862: lii) was apparently the first author to recognise the subfamily Empidinae , and also the first to arrange the empidids into five subfamilies (from Chvála 1983: 54). This subfamily is defined by the following suite of characters: palpi normally strongly upcurved, broad wings (anal lobe usually developed), forelegs not raptorial, empodium usually bristleform. Cumming (1994) hypothesized that presentation of nuptial gifts to females by males, correlated with loss of hunting ability in females, might represent two additional synapomorphies of the Empidinae (or at least a portion of the subfamily), although mating behaviors of the more plesiomorphic genera, such as Edenophorus , Empidadelpha and Sphicosa , are presently unknown. The Empidinae includes the following tribes:
Collin (1961: 326) was perhaps the first worker to separate the Empididae into two tribes. This tribe is presently undefined and undoubtedly paraphyletic and presently simply a dumping group for all genera not assigned to the Hilarini . This lineage is distinguished from Hilarini on the basis of a setose laterotergite, cercus enlarged and functioning as a clasping organ, costa usually ending at or just beyond R 4+5, vein R 1 of constant thickness and male tarsomere 1 of foreleg rarely enlarged ( Bickel 1996a). The Empidini includes the following genera: Clinorhampha Collin , Empidadelpha Collin , Empis Linnaeus , Hystrichonotus Collin , Lamprempis Wheeler and Melander , Macrostomus Wiedemann , Opeatocerata Melander , Porphyrochroa Melander , Rhamphella Malloch , Rhamphomyia Meigen , and Sphicosa Philippi ( Smith 1980; Chvála & Wagner 1989; Rafael & Cumming 2004).
Edenophorus Smith View in CoL was originally assigned to the Microphorinae View in CoL when male specimens were not known. It was later transferred to the Ocydromiinae ( Chvála, 1981) View in CoL , but males still remained unknown. Ulrich (1994) believed it showed affinities to the Empididae sensu Chvála (1983) View in CoL . Male specimens were identified by Sinclair (2002), confirming its assignment to the Empidinae View in CoL . However, on the basis of the absence of strongly arched palpi, Edenophorus View in CoL is excluded from the clade comprising Empidadelpha View in CoL , Empidini View in CoL and Hilarini ( Sinclair 2002) View in CoL . It is tentatively assigned herein to the Empidini View in CoL until a better definition or diagnosis of this tribe is made available.
Hilarini Collin, 1961: 326 View in CoL . Type genus Hilara Meigen, 1822 View in CoL .
The tribe is well defined on basis of laterotergite usually bare, cercus usually small and desclerotised, hypandrium usually forming a keellike hood over the phallus along the posterior margin, costa usually circumambient, R 1 thickened before it joins the costa and male tarsomere 1 of foreleg enlarged ( Bickel 1996a). The following genera are included in the Hilarini View in CoL : Afroempis Smith View in CoL , Allochrotus Collin View in CoL , Amictoides Bezzi View in CoL , Aplomera Macquart View in CoL , Atrichopleura Bezzi View in CoL , Bandella Bickel View in CoL , Cunomyia Bickel View in CoL , Deuteragonista Philippi View in CoL , Gynatoma Collin View in CoL , Hilara Meigen View in CoL , Hilarempis Bezzi View in CoL , Hilarigona Collin View in CoL , Hybomyia Plant View in CoL , Pasitrichotus Collin View in CoL , Thinempis Bickel View in CoL , and Trichohilara Collin View in CoL ( Smith 1980; Cumming 1994; Bickel 1996a, 1998, 2002).
Bickel (1998) considered Cunomyia primitive with respect to other Hilarini on the basis of the form of its male cercus and postabdominal structure. The male eyes are separated on the frons in this genus and the upper facets are not enlarged. It is possibly closely related to Bandella ( Bickel 2002) . Members of the Hilara flavipes Meigen complex have holoptic males with the upper ommatidia enlarged and Chvála (1999) has considered this the ground plan state based on outgroup comparison with the Empidini . This head morphology is co–related to primitive unmodified small synorchesic swarms.
Hemerodrominae Schiner, 1862 : li, lii. Type genus Hemerodromia Meigen, 1822 View in CoL .
Hemerodromiinae Wheeler & Melander, 1901: 376 .
This is one of the most distinctive empidoid subfamilies, characterized by raptorial
forelegs well separated from the midlegs and the fore femur bearing at least a single
ventral row of stout setae. Some genera possess aquatic larvae, with adults of many genera
found on emergent rocks and riparian vegetation of streams and rivers. The larva of
Chelipoda is terrestrial ( Trehen 1969). The Hemerodromiinae is divided into two tribes.
Chelipodini Hendel, 1936: 1929 View in CoL .
This tribe can be separated from Hemerodromiini View in CoL on the basis of the long arista more than twice the length of the postpedicel, scutum with welldeveloped setae, laterotergite with setae and male terminalia reflexed over abdomen ( MacDonald 1988). The following genera are included in the Chelipodini View in CoL : Afrodromia Smith View in CoL , Chelipoda Macquart View in CoL , Chelipodozus Collin View in CoL , Drymodromia Becker View in CoL , Monodromia Collin View in CoL , Phyllodromia Zetterstedt View in CoL , and Ptilophyllodromia Bezzi View in CoL ( Smith 1967, 1975, 1980, 1989).
The genus Cephalodromia Becker is not an empidoid and is assigned to the Mythicomyiidae (Platypyginae) (see Greathead & Evenhuis 2001). However, the generic assignment of the Taiwan species, C. pictipennis Bezzi , requires examination, but it should probably be tentatively assigned to Chelipoda until the Chelipodini can be defined on a world basis.
This tribe can be separated from Chelipodini on the basis of the short antennal style less than twice the length of the postpedicel, scutum lacking well developed setae, laterotergite bare and male terminalia erect or projecting posteriorly ( MacDonald 1988). The following genera are included in the Hemerodromiini : Chelifera Macquart , Cladodromia Bezzi , Colabris Melander , Doliodromia Collin , Hemerodromia Meigen , Metachela Coquillett and Neoplasta Coquillett ( Smith 1967, 1975, 1980, 1989).
Clinocerinae Collin, 1928: 2 View in CoL , 3, 59. Type genus Clinocera Meigen, 1803 View in CoL .
Clinoceratinae Melander, 1928: 10 .
The Clinocerinae View in CoL is a very well defined lineage and one of the few lineages where the phylogenetic relationships of the genera have been evaluated with modern techniques ( Sinclair 1995a). Anumber of genera remain to be described from the Australasian region. The larval habitat of the majority of taxa is considered aquatic, normally streams and rivers with rocky substrates. The most primitive genera dominate the Southern Hemisphere. The subfamily can be identified by the stout, erect costal setae, suckerlike labellum, pulvilliform empodium, anal lobe of wing not developed and CuA 2 recurved at apex of cell cu p ( Sinclair 1995a). The following genera are included in the Clinocerinae View in CoL : Aclinocera Yang & Yang View in CoL , Afroclinocera Sinclair View in CoL , Asymphyloptera Collin View in CoL , Bergenstammia Mik View in CoL , Clinocera Meigen View in CoL , Clinocerella Engel , Dolichocephala Macquart View in CoL , Hypenella Collin View in CoL , Kowarzia Mik View in CoL , Oreothalia Melander View in CoL , Phaeobalia Mik , Proagomyia Collin View in CoL , Proclinopyga Melander View in CoL , Rhyacodromia Saigusa View in CoL , Roederiodes Coquillett View in CoL , Trichoclinocera Collin View in CoL , and Wiedemannia Zetterstedt ( Sinclair 1995a).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Empididae
| Sinclair, Bradley J. & Cumming, Jeffrey M. 2006 |
Hilarini
| Collin, J. E. 1961: 326 |
Chelipodini
| Hendel, F. 1936: 1929 |
Clinocerinae
| Collin, J. E. 1928: 2 |
Clinoceratinae
| Melander, A. L. 1928: 10 |
Hemerodromiinae
| Wheeler, W. M. & Melander, A. L. 1901: 376 |
Empididae
| Giebel, C. G. 1856: 206 |