
SCIADOCERINAE, Schmitz, 1929
|
publication ID |
https://doi.org/10.1111/zoj.12208 |
|
persistent identifier |
https://treatment.plazi.org/id/039987A3-BD52-297F-6561-FA3528508A4E |
|
treatment provided by |
Felipe |
|
scientific name |
SCIADOCERINAE |
| status |
|
SUBFAMILY SCIADOCERINAE View in CoL
No thorax characters were found to support this grouping, although some of the previous characters known in the literature (see characters 59–61, below) point to the monophyly of the subfamily.
Euphorida (unranked) = non- sciadocerine phorids
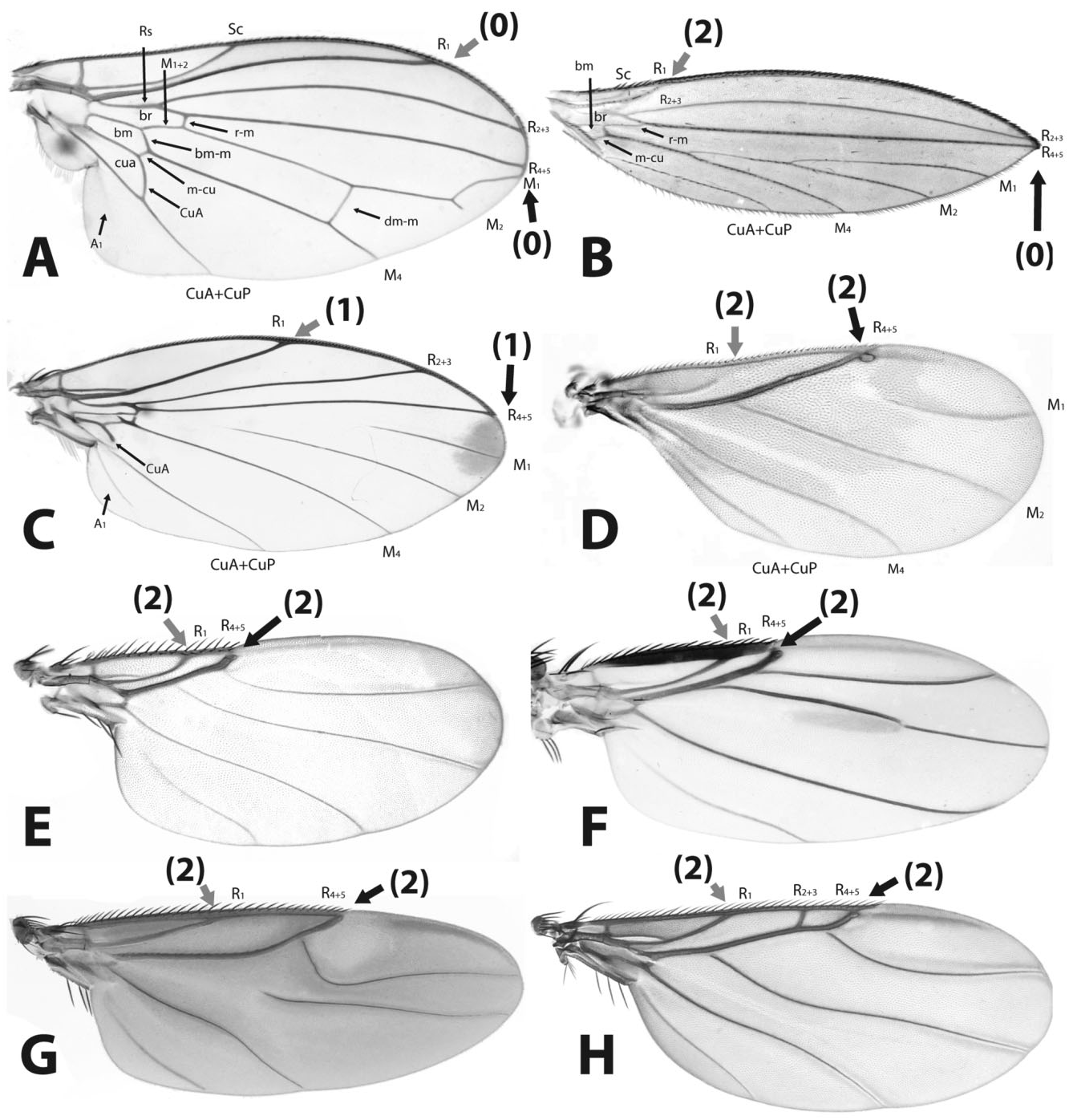
9. Vein M 2: (0) basally terminating posterior to R 4+5, (1) appearing to be attached to R 4+5 ( Fig. 13A–H View Figure 13 ).
McAlpine & Martin (1966) noted that the base of M was lost, but did not consider these veins further. The condition in Latiborophaga ( Fig. 13G View Figure 13 ) is obviously a case of reversal within the Phorinae . 10. Costal setae: (0) short, thick, spine-like, (1) elongate, thin, (2) elongate, thick, feathered ( Fig. 17A–I View Figure 17 ).
Hackman & Väisänen (1985) reviewed the costal setation of Diptera , providing a classification of the setal patterns. Disney (1988) proposed a new character state for Ironomyiidae + Sciadoceridae , indicating they were not necessarily closely related to Phoridae . Brown (1992) examined several Platypezidae and found that their setation was the same as that depicted by Disney (and thus plesiomorphic) ( Fig. 17A, B, D, E View Figure 17 ). The Lonchopteridae have longer, thin setae up to the end of vein R 1 ( Fig. 17C View Figure 17 ), somewhat similar to the condition seen in higher phorids, whose longer setae extend along the entire length of the costa. In Phoridae , the costal setae are elongate, with those of Chonocephalus , Cyphocephalus and termitoxeniines being thin and hair-like (state 1, Fig. 17F, G View Figure 17 ), and those of Metopininae and Phorinae being thicker, bristle-like and feathered (state 2, Fig. 17H, I View Figure 17 ; feathering difficult to see in Fig. 17H View Figure 17 ). The setae of Chonocephalus and Cyphocephalus are relatively shorter than those of other euphorids, possibly indicating another character state (costal setae longer) supporting the grouping of termitoxeniines with Phorinae + Metopininae .
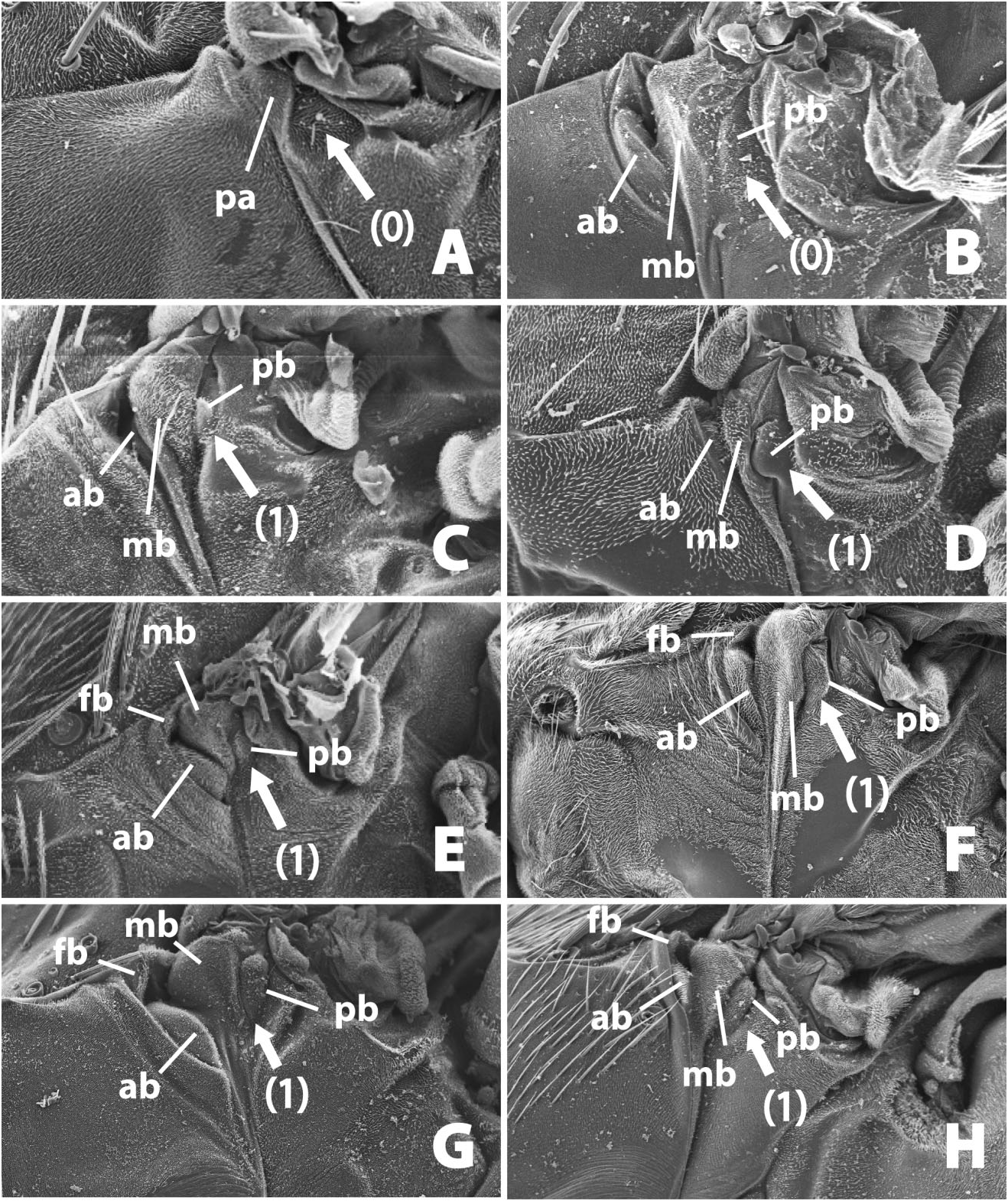
11. Anterodorsal extension of anepimeron: (0) flat, (1) with posterior bump ( Fig. 18A–H View Figure 18 ).
This area of the thorax is discussed above, in the overview of the evolution of thoracic structure. This posterior bump is similar in location to the greater ampulla in calyptrate Diptera . 12. Angle of mesopleural suture meeting the anapleural suture: (0) close to 90°, (1) at an obtuse angle in relation to the anapleural suture ( Fig. 19A–F View Figure 19 ).
The angle that the mesopleural suture meets the anapleural suture is related to the relative reduction in size of the anepimeron and the development of the anepisternum posterodorsally. The Platypezidae and the Lonchopteridae retain the plesiomorphic state, also seen in other brachyceran families ( Fig. 19A View Figure 19 ). Within the phorids, the apomorphic condition is the mesopleural suture not being at a right angle ( Fig. 19B–F View Figure 19 ).
13. Meron: (0) relatively small, equal in height and width, (1) enlarged, much higher than wide ( Fig. 20A–F View Figure 20 ).
An enlarged meron, much higher than wide, is characteristic of all Euphorida. This sclerite, however, corresponds to the meron entirely fused with the katepimeron. A narrow katepimeron is still visible in other brachyceran families (e.g. Therevidae , Fig. 1 View Figure 1 ) and Platypezidae ( Figs 2, 20A View Figure 20 ). In Lonchopteridae ( Fig. 3) and in Sciadocerinae ( Figs 4 View Figure 4 , 20B View Figure 20 ) not even a depression between the katepimeron and the meron can be identified. In all these taxa, however, whether a suture is present or not, the shape of the plate is approximately the same ( Fig. 20A, B View Figure 20 ). The shape of the plate in Chonocephalus ( Fig. 20C View Figure 20 ) is clearly apomorphic in relation to that of Sciadocera , as is seen in metopinines and phorines ( Fig. 20D–F View Figure 20 ). In a few euphorids, as in Phlebothrix , there are other modifications of the shape of the posterior part of the thorax, so the meron may be shorter.
14. Katepisternum: (0) pruinose, (1) mostly bare ( Fig. 21A–F View Figure 21 ).
In platypezids ( Fig. 2), lonchopterids ( Fig. 3, 21A View Figure 21 ) and sciadocerines, nearly the entire pleuron is pruinose (covered with microtrichia). In phorids, at least the lower two-thirds of the katepisternum lacks microtrichia. The condition in Chonocephalus ( Fig. 21B View Figure 21 ) is relatively apomorphic in relation to that in sciadocerines, but the katepisternum still has an extensive microtrichia cover at the dorsal and posterior margins. Perhaps an additional character state could be proposed for the condition seen in Chonocephalinae , but for the time being we decided to keep only two states. The termitoxeniines, metopinines and phorines have the katepisternum typically bare and shining ( Fig. 21C–F View Figure 21 ).
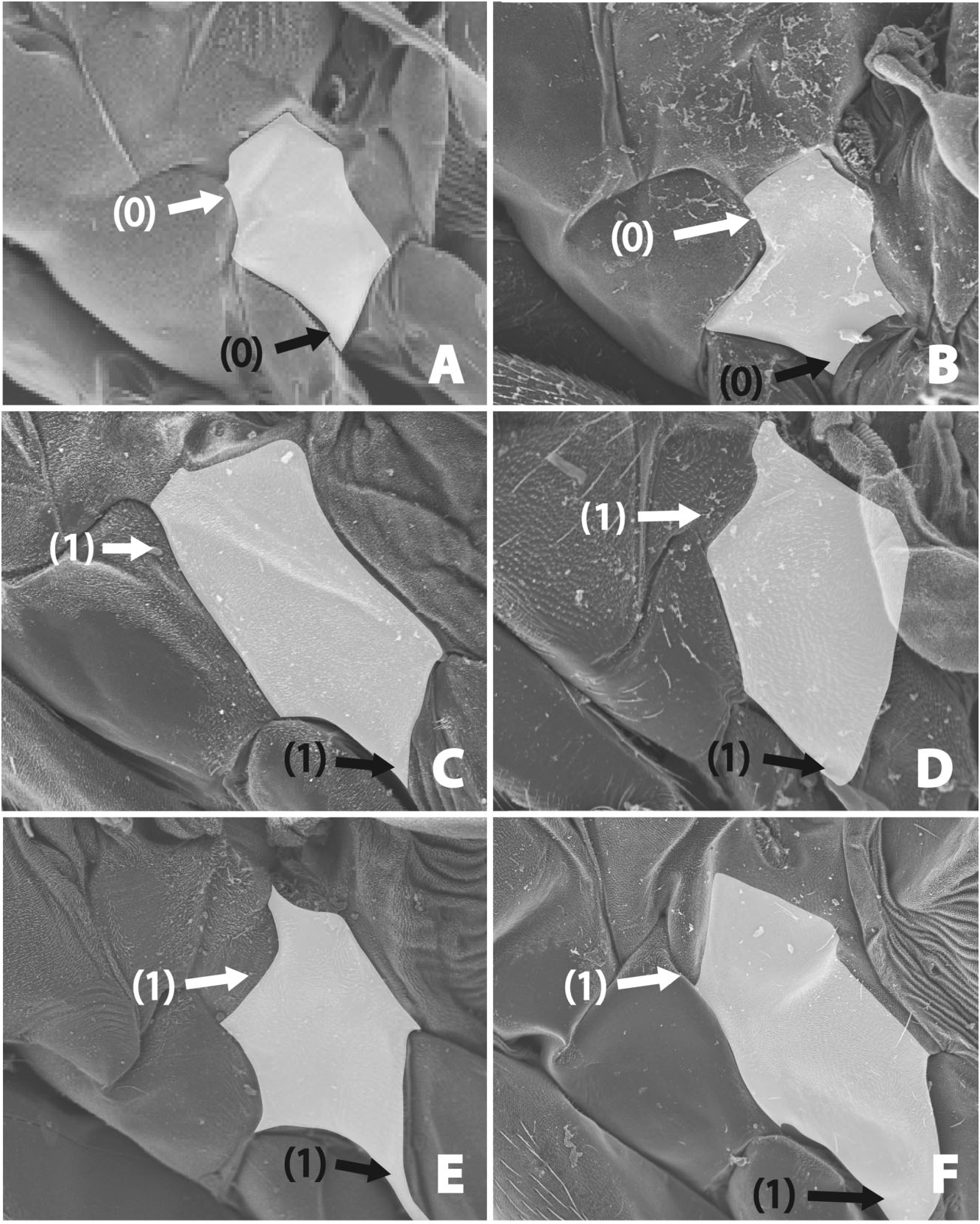
15. Anterior spiracle: (0) ovoid or reniform, (1) small, round, (2) large, round ( Fig. 22A–F View Figure 22 ).
The shape of the anterior spiracle is highly diagnostic of each group. Outgroups, including Platypezidae ( Figs 1 View Figure 1 , 22A View Figure 22 ) and Lonchopteridae ( Fig. 3), as well as Sciadocerinae ( Figs 4 View Figure 4 , 22B View Figure 22 ), have a narrow ovalshaped spiracle, similar to that in at least some nonbrachyceran Diptera (such as Empididae and Dolichopodidae ). A round but small spiracle is found in Chonocephalus ( Fig. 22C View Figure 22 ) and termitoxeniines ( Fig. 22D View Figure 22 ). Unexpectedly, Aenigmatias and some thaumatoxenines also have relatively small anterior spiracle, possibly as an adaptation to living in social insect nests. Other phorids have a large round anterior spiracle ( Fig. 22E, F View Figure 22 ).
16. Posterior margin of anepisternum appearing: (0) ‘peeled back’, (1) folded partly over anterior bump ( Fig. 23A–F View Figure 23 ). This area of the thorax is discussed above, in the overview of the evolution of thoracic structure.
17. Lateral parapsidal suture (suture between scutum and pleuron) and lateral ridge of scutum: (0) straight, extended anteriorly over spiracle, (1) with only short vestige of carina ending near to spiracle, (2) stopping well posterior of anterior spiracle and joining pronotal suture ( Fig. 24A–F View Figure 24 ).
A paratergite is not characterized in the Platypezoidea, so the lateral parapsidal suture and the lateral ridge of the scutum cannot be properly distinguished in two separate structures. In platypezoids there is a fusion between scutum, postpronotum, proepisternum and anepisternal sclerotization around the anterior spiracle. In Platypezidae (and as well in lonchopterids and sciadocerines, Fig. 24A View Figure 24 ), the lateral parapsidal suture clearly reaches anterior to the anterior spiracle. The suture is slightly shorter in Chonocephalus ( Fig. 24B View Figure 24 ) and the termitoxeniines ( Fig. 24C View Figure 24 ), but a carina still surrounds the spiracle. In some phorines and metopinines ( Fig. 24D View Figure 24 ) the carina stops near to the spiracle, although in at least a few metopinine genera, such as Apodicrania , an even shorter suture can be seen. In some phorines, the lateral parapsidal suture itself ends well posterior to the anterior spiracle ( Fig. 24F View Figure 24 ). This character defines a newly recognized grouping of the Borophaga group of genera (part of Brown’s 1992 concept of Hypocerinae ), the diplonevrines (= Aenigmatiinae of Brown, 1992, minus Aenigmatias ) and three genera formerly considered part of the genus Chaetopleurophora . All exhibit the same shortening of the lateral parapsidal suture with its dorsal apex curved strongly posteriorly and joining the pronotal suture.
18. Metepimeron: (0) sclerotized, recognizable, broad, (1) reduced to at most narrow edge along posteri- or margin of meron ( Fig. 25A–F View Figure 25 ).
The plesiomorphic condition of this sclerite can be seen in most schizophoran families ( Figs 1–3 View Figure 1 , 25A View Figure 25 ), in which the metepimeron is considerably smaller than other sclerites, but is present. Within the phorids, the sciadocerines have a considerably well-developed metepimeron ( Figs 4 View Figure 4 , 25B View Figure 25 ). In the euphorids, including the chonocephalines ( Fig. 25C View Figure 25 ), the metepimeron is sometimes hardly recognizable as an independent sclerite, with only the suture posterior to the metepimeron being visible ( Fig. 25D–F View Figure 25 ). In some cases, a slender sclerite can be seen.
19. Halter and posterior spiracle position: (0) closer to midpoint between wing base and dorsal margin of hind coxa, (1) close to wing base ( Fig. 26A–F View Figure 26 ).
The position of the halter and the posterior spiracle in sciadocerines ( Figs 4 View Figure 4 , 26B View Figure 26 ) is similar to that seen in platypezids ( Fig. 26A View Figure 26 ) and other brachycerans ( Figs 1–3 View Figure 1 ), which is at about half of the pleural height. In sciadocerines the posterior spiracle is slightly more ventral than the anapleural suture. This condition is modified in Metopininae + Phorinae ( Fig. 26E, F View Figure 26 ), with the displacement in the halter and the posterior spiracle to a more dorsal position of the thorax. In Chonocephalus ( Fig. 26C View Figure 26 ), it is clear that the ventral half of the thorax is greatly enlarged relative to the dorsal half, but the position of the posterior spiracle and the halter is still not particularly displaced dorsally, compared with the anapleural suture. These changes may be connected with the development of the walking musculature in the ventral half of the thorax, and a reduction of the size of the posterodorsal part of the thorax, especially the laterotergite and mediotergite, which are much smaller in euphorids than in most brachycerans.
20. Katepisternum: (0) straight across top, (1) projected into anepimeron ( Fig. 27A–F View Figure 27 ).
This character is responsible for the development of the W-shape of the anepimeron in most phorids. The posterodorsal extension of the katepisternum results in the W-shape of the anepimeron in a large clade in the family. The katepisternum in other brachycerans ( Figs 1–2 View Figure 1 , 27A View Figure 27 ) is slightly projected posteriorly beyond the mesopleural suture, in such a way that the posterior half of the katepisternum is ventral to part of the anepimeron. In these groups, the dorsal margin of the anapleural suture is more or less straight. This plesiomorphic condition is well preserved in the sciadocerines ( Fig. 4 View Figure 4 ), in which the posterior end of the katepisternum is slightly projected dorsally into the anepimeron. Lonchoptera ( Fig. 3) has a relatively apomorphic condition. In Chonocephalus ( Fig. 27B View Figure 27 ) there is some projection of the katepisternum posterodorsal corner, which in metopinines and phorines is deeper in different degrees ( Fig. 27D–F View Figure 27 ). Different changes in other pleural sclerites strongly modify the condition seen in these subfamilies and there is a wide variety of shapes of the anepimeron. We still decided to not code most of the variation in the shape of the anepimeron found in the metopinines and phorines.











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.