
Tyrannosaurus, Osborn, 1905
|
publication ID |
https://doi.org/ 10.1111/j.1095-8312.1970.tb01707.x |
|
DOI |
https://doi.org/10.5281/zenodo.3680526 |
|
persistent identifier |
https://treatment.plazi.org/id/03996806-FFCF-FFE6-8178-F9BE29FDFEA9 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus |
| status |
|
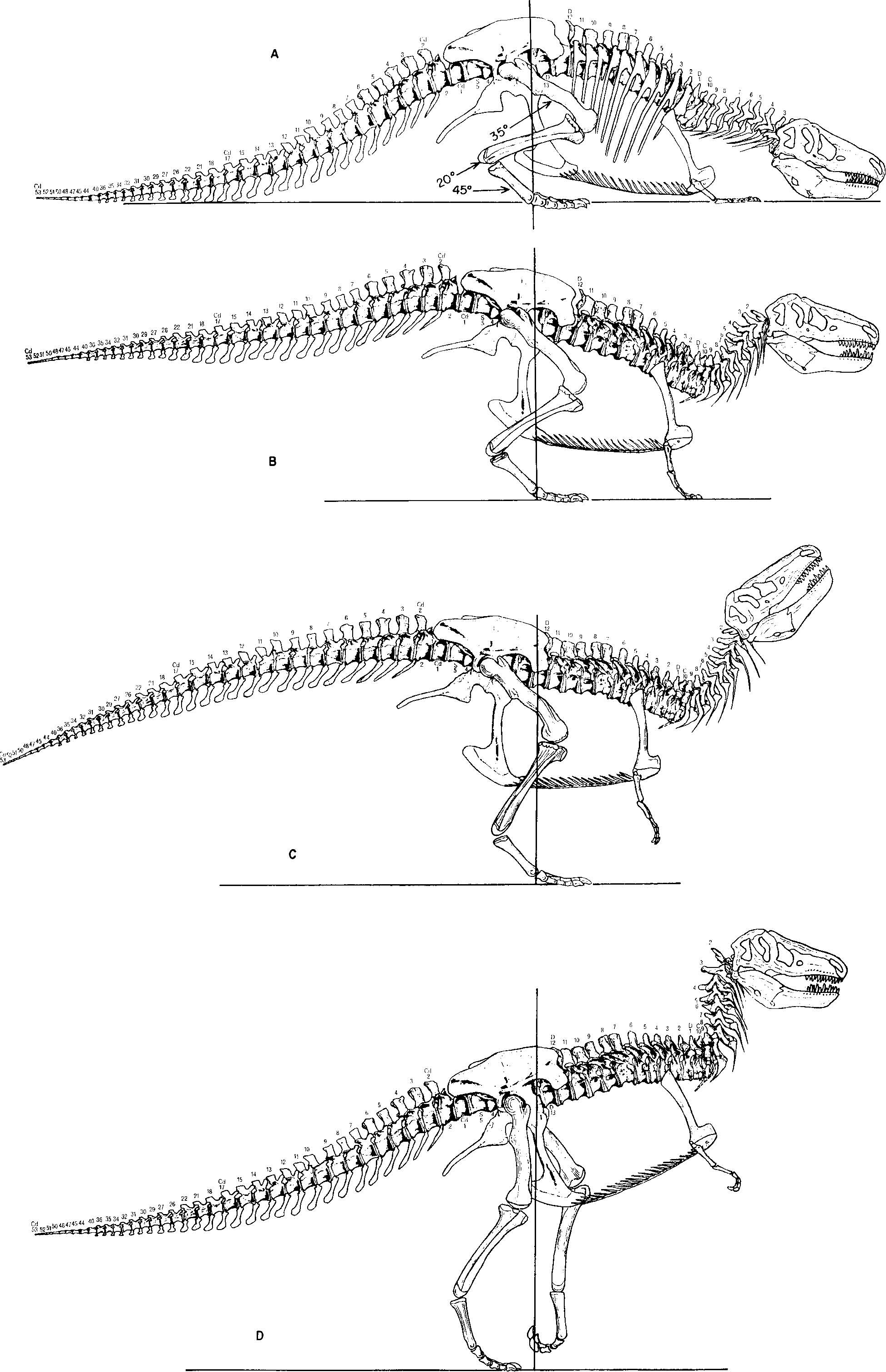
Tyrannosaurus is the flesh-eating dinosaur which is perhaps best known to the general public in the form of restorations based on the composite skeleton mounted in the American Museum of Natural History. This shows the animal with the distal part of the tail resting on the ground, the proximal part of the tail and the dorsal vertebral column forming a straight line at an angle of about 45 " with the ground ( Romer, 1966). Until now the stance of the mounted specimen in New York View Materials has been accepted uncritically and there have been no significant departures from this classic pose, with the exception of reconstructions by Augusta & Burian (1958) which shows the animal with its tail off the ground. Recently the British Museum (Natural History) purchased the partial skeletons of two View Materials individuals View Materials which, with the missing parts reconstructed, make up the composite skeleton now displayed in the Dinosaur Gallery. One purpose of this paper is to outline the reasons for the marked departures from previously accepted restorations made in preparing the B.M. (N.H.) exhibit. The following aspects of this skeletal reconstruction and its posture are contested: the length of the tail, the angle of the vertebral column, and the positioning of the limbs and skull.
VERTEBRAL COLUMN AND SKULL
The best-preserved vertebral column was figured by Osborn (1917). The 7th and 8th dorsal vertebrae have their centra fused, the neural spines of the 7th and 8th and the 9 th- 12 th almost touch, and there is evidence of strong connective tissue linking the neural spines of all the dorsal region vertebrae. Hence this part of the dorsal region of the column must have been rigid. If the skull were articulated on the vertebral column as restored by Osborn the occipitalcondylewould not have been in full articulation with the atlas. In fact, the skull was supported on a well-defined cervical region and at a marked angle to the rest of the column ( Fig. 1 D View FIGURE 1 ). If the dorsal region is orientated horizontally, the 'swan-neck' would permit the skull to be carried horizontally and the flexible nature of the neck would also enable the animal to manoeuvre its head and reach the ground, an important factor when feeding.
TAIL
With the dorsal part of the vertebral column held in the horizontal plane, the sacrum would have pivoted forwards,at the same time bringing the tail free of the ground to act as a counter-balance to the trunk. The tail as previously restored is exceptionally long when compared with that known from the closely related Gorgosaurus . Since only 20 caudal vertebrae of the New York Tyrannosaurus are authentic, it would be reasonable to postulate a tail comparable to that of Gorgosaurus, ( Lambe, 1917) . If this is accepted, then some 12feet of the tail of the New York specimen must be removed.
Haemal arches on the caudal vertebrae have been taken as evidence that the tail was dragged along the ground, and are alleged to have had a protective function. Their presence as far forward as the second and third caudal vertebrae makes this difficult to accept, since it is hardly likely that the proximal part of the tail made contact with the ground and thus protection from abrasion was unnecessary. Moreover, the footprints of carnivorous dinosaurs give little indication that the tail was dragged along the ground. Although the American restoration indicates a very flexible tail, with appropriate short zygopophysesfor the distal caudal vertebrae, in the comparable Gorgosaurus , where the distal region of the tail is preserved, the zygapophyses are extremely long, indicating a severe restriction on the lateral movement of the distal end of the tail, ( Lambe, 1917).
HIND-LIMB
To work out the functioning of the hind-limb it is useful to consider the living flightless birds, where the femur is shorter than the tibia and is held forward of the acetabulum. As there is no tail to act as a counter-balance, this orientation of the femur serves to bring forward the centre of gravity. In Tyrannosaurus the femur is longer than the tibia and the limb could not have borne the stresses if it were held horizontally. In fact, the presence of the heavy tail would have brought back the centre of gravity of the animal and made the horizontal position of the femur unnecessary. When walking, the weight of the animal must have been borne by one leg at a time alternately, and in birds and carnivorous bipedal dinosaurs comparable specializations are found. In both groups the major axis through the articular surface of the proximal end of the femur is at a marked angle to that of the distal end. When the animal supports its weight on one leg, the distal articular surface of the femur rolls out laterally, thus tightening the ligament between the great trochanter and the cnemial crest. This results in the kneejoint being moved laterally outwards and the animal’s weight being transferred sideways so that the centre of gravity lies over the limb, the tightened ligament preventing excess dexure. In Tyrannosaurus , as the free limb swung forwards the tail would have swung laterally away from it, whilst the femur would be drawn through a restricted arc and the main part of the stride accomplished by the movement at the knee-joint. The large procnemial crest on the anterior surface of the proximal part of the tibia would be suitable for the insertion of tendonous tissue for the attachment of muscles providing leverage for pulling the lower part of the limb forwards. As the limb was bent and the ankle joint flexed, the tendons along the posterior surface of the limb connecting the tibia and fibula to the more distal elements would have been stretched, resulting in the clenching of the toes as in birds. This would allow the foot to clear the ground as the limb swung forwards.
Since the weight must have been carried by each limb in turn, the gait is likely to have been sinuous, producing a bird-like waddling, and the series of megalosaurian footprints from the Purbeck Beds (Upper Jurassic) of Swanage now on display in the Dinosaur Gallery at the British Museum (Natural History) confirm this view. The individual footprints do not lie in a straight line and the toes point inwards, indicating that the animal was pigeon-toed. The stride was small (some 25 1/2 inches) in relation to the size of the animal's foot (12 inches). A detailed map of the trackway reveals an overall sinuous curve and seems to confirm the essentially bird-like gait of these animals, whilst the shortness of the stride further confirms the restricted amount of posteroanterior movement at the acetabulum. Colbert (1965) restores Antrodemus (Allosaurus) and Gorgosaurus in exactly the posture here suggested for Tyrannosaurus , although he does not give his reasons for so doing.
Although it has long been recognized that the so-called 'vestigial' fore-limbs of Tyrannosaurus were too small to have played any part in locomotion (see, for example, Romer 1966) the scapulae and coracoids are large and must have been associated with a considerable pectoral musculature. The humeri although short are massive ( Osborn, 1906) and, allowing for the muscles and skin, the width across the humeral region would have been compared to that of a human thigh. The fore-limb would have been correspondingly muscular. With the large muscular pectoral girdle to support it, the fore-limb would certainly have been powerful enough to raise the chest region off the ground.
When in a position of rest, the hind-limbs would be folded under the body in much the same way as a hen's, with the fore-limbs in contact with the ground and the skull extended so that the lower jaw rested on the ground ( Fig. 1 A View FIGURE 1 ). With the hind-limbs folded, each femur would have been held at approximately 35 " to the horizontal, with the tibia bent back at about 20" and the metatarsals bent forwards at 45 ".
I n order to rise, the animal had to extend its hind-limbs, but the effect of beginning such an action would be to push the body forwards along the ground. The role of the fore-limbs was that of a brake holding the body, so that the force exerted by the extension of the hind-limbs was transmitted to the pelvic region, thus pushing it upwards. The fore-limbs would have assisted in the initial stages of lifting the trunk region off the ground.
From a resting position the animal's first action in rising would be to raise the head and at the same time begin to straighten the fore-limbs ( Fig. 1 A View FIGURE 1 ).Next the hind-limbs began to extend, the skull was raised higher, and the fore-limbs were further extended, while the tail was lifted off the ground ( Fig. 1 B View FIGURE 1 ).At the next stage the animal must have continued straightening the fore-limbs until the trunk region was pressed upwards to their full extent; at the same time the head was thrown upwards and backwards, whilst the hind-limbs continued to extend and the tail was depressed ( Fig. 1 C View FIGURE 1 ). The muscles from the ventral and lateral caudal regions and the ischia, acting on the femora, served to assist the extension of the hind-limbs and helped to maintain equilibrium, the centre of gravity being maintained at a line through the ankle and knee-joints and the anterior end of the sacrum. Figure 1 D View FIGURE 1 shows the hind-limbs fully extended, and the head and tail carried in the normal position of equilibrium.
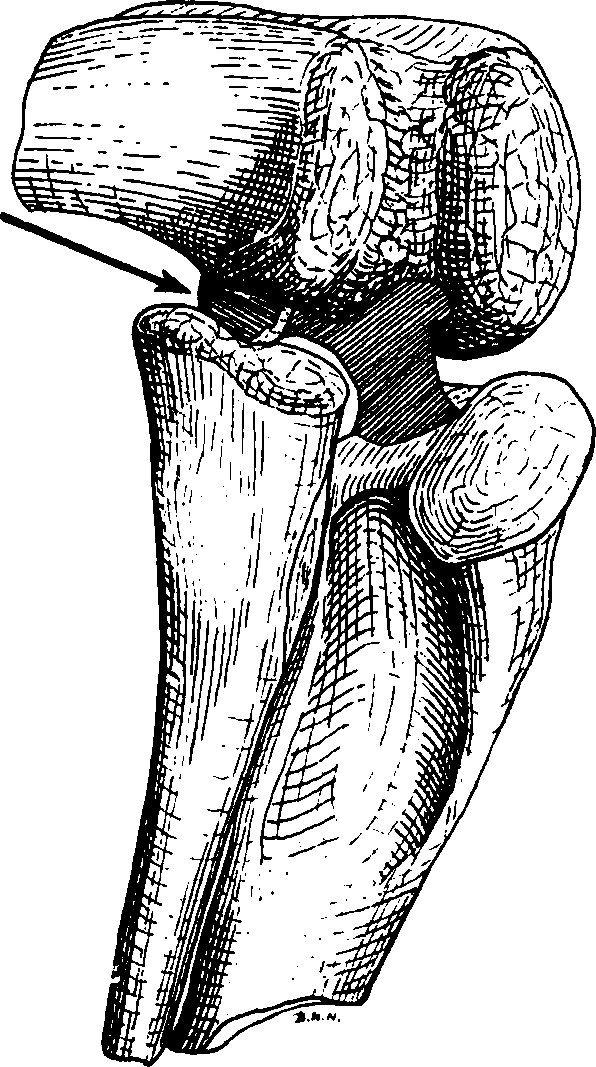
Owing to the acute angles adopted by the femorae and tibiae when the animal was at rest, there must have been considerable lateral forces acting on the knee-joints and tending to force them outwards, in which case there would have been great difficulty in flexing the legs. However, any tendency for a knee-joint to be forced outwards was prevented by a condyle, situated behind the outer articular surface of the distal end of the femur, this occupied the space between the fibula and tibia when the knee was fully bent, as shown in Fig. 2 View FIGURE2 .
CONCLUSION
The appearance in life of Tyrannosaurus must have been quite different from that previously imagined and Fig. 1 D View FIGURE 1 shows the new restoration, which emphasizes the bird-like stance. From a consideration of the skeleton, as well as the fossilized tracks of related dinosaurs, it appears that the gait was an ungainly waddling rather than the formerly postulated majestic striding.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |