
Geosesarma spectrum, Ng & Ng, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4614.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:2A3C63DE-4C89-4433-80F8-FD5FFC250DDA |
|
persistent identifier |
https://treatment.plazi.org/id/0394AC51-9C78-FF88-748F-FF40FDC7FBC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Geosesarma spectrum |
| status |
sp. nov. |
Geosesarma spectrum View in CoL n. sp.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 5–10 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Geosesarma cf. gracillima — Choy, 1991: 129 , 144 (not Sesarma (Sesarma) ocypoda gracillima De Man, 1902 View in CoL )
Material examined. Holotype: male (13.6 × 13.5 mm) (ZRC 2018.0740), forest trails around Ulu Ulu resort, Ulu Temburong National Park, 4°33’21.2”N, 115°9’16.1”E, Brunei Darussalam, coll. P.Y.C. Ng, September 2018. Para- types: 2 males (7.9 × 7.6 mm, 12.2 × 12.1 mm), 2 females (15.1 × 14.3 mm, 10.2 × 9.8 mm), 1 ex-ovigerous female with 10 juvenile crabs (11.2 × 10.7 mm) (ZRC 2018.0741), same data as holotype. Others: 1 young male (ZRC 2018.0744), on leaf of sapling, waist height, trail to West Ridge, Belalong, Brunei, coll. I. Das, 27 March 1992; 1 male (10.8 × 10.7 mm) (ZRC 2018.0745), Kuala Belalong Field Studies Centre, Brunei, coll. S. Choy, 10 February 1991; 2 males (10.8 × 10.7 mm, 10.2 × 9.8 mm), 2 females (10.5 × 10.0 mm, 10.2 × 9.9 mm) (ZRC 2017.1215), in pitfall trap, West Ridge, Kuala Belalong Field Studies Centre, Brunei, coll. J. K. Charles, 7 February 1993; 2 males (larger 10.8 × 10.5 mm), 1 female (ZRC 2011.1023), station 19, near Pagon House, on forest floor, Kuala Belalong Field Studies Centre, Brunei, coll. Samahan, 15 February 1993; 1 young male (5.7 × 5.4 mm) (ZRC 2018.0746), in Nepenthes bicalcarata (Hooker, 1893: Nepentheceae), Seria, Brunei, coll. C. Clarke, 29 August 1989; 1 young male (ZRC 2016.0135), Belait peat swamp, Sungei Mendaram, Brunei, coll. I. Kwan, 18 February 2013; 1 juvenile female (ZRC 2018.0747), Lamunin, Brunei, coll. 30 October 1989. Comparative material. Geosesarma katibas, Ng, 1995a : holotype male (13.5 × 13.5 mm) ( ZRC 1996.01834), among dead leaves, quadrat about 50 m from Sungei Katibas, east bank, about 100 m from mouth of Melinau, Ulu Katibas, Song District, Sarawak, Malaysia, coll. R.B. Stuebing, 25 March 1994; 1 paratype male (13.2 × 13.1 mm) ( ZRC 1995.0274 View Materials ), same data as holotype ; 3 males (largest 12.2 × 11.9 mm), 2 females (largest 12.3 × 11.8 mm) ( ZRC 2016.0296 View Materials ), Sungei Boeng, tributary of Sungei Julau, Sarikei, Sarawak, Malaysia, coll. J. Grinang, 2 January 2013 ; 1 young male ( ZRC 2018.0748 View Materials ), station Q99, East bank, Sungei Katibas, Sarawak, coll. 22 March 1994 ; 1 male (carapace crushed, ca. 14 mm carapace width) ( ZRC 2007.0257 View Materials ), on road from Samarahan to Bintulu Division, coll. T.M. Leong, 14 April 2007 .
Geosesarma gracillimum (De Man, 1902) : 6 males (largest 12.1 × 11.8 mm), 7 females (largest 11.1 × 10.4 mm), 1 ovigerous female (12.0 × 11.6 mm) (egg size 1.2 mm in diameter after preservation) (ZRC 2007.262), besides trail in Glen Forest, arboreal, Lambir Hills National Park, Sarawak, Malaysia, coll. T.M. Leong et al., 22, 23, 25 April 2007; 1 female ( ZRC 2011.1031 View Materials ), on leaf, far from water, near termite mound, Lambir Hills National Park , Sarawak , coll. H.S. Yong, 1995; 1 male (13.0 × 12.8 mm) ( ZRC 2011.1030 View Materials ), Niah National Park , Sarawak, coll. local rangers, 19 February 2001 .
Geosesarma bau Ng & Grinang, 2004: 1 View in CoL female (ZRC 2018.0750), in cup of Nepenthes ampullaria ( Jack, 1835) [Nepentheceae], Mount Santubong, Sarawak, Malaysia, coll. W. Denzer, 10 April 1992; 1 female (9.6 × 9.3 mm) ( ZRC 2002.0228 View Materials ), road from Kuching to Serian, 10th mile rock, into Kuap, 1°25’8.9” N 110°20’52.9”E, Sarawak, Malaysia, coll. P.K.L. Ng et al., 5 September 1995 GoogleMaps ; 1 ovigerous female (11.2 × 10.9 mm) (egg size about 1.0 mm in diameter, eggs wrinkled) ( ZRC 1965.7.29.5), River Chuat , Sibu, near Sarikei, Sarawak, Malaysia, coll. 1948 .
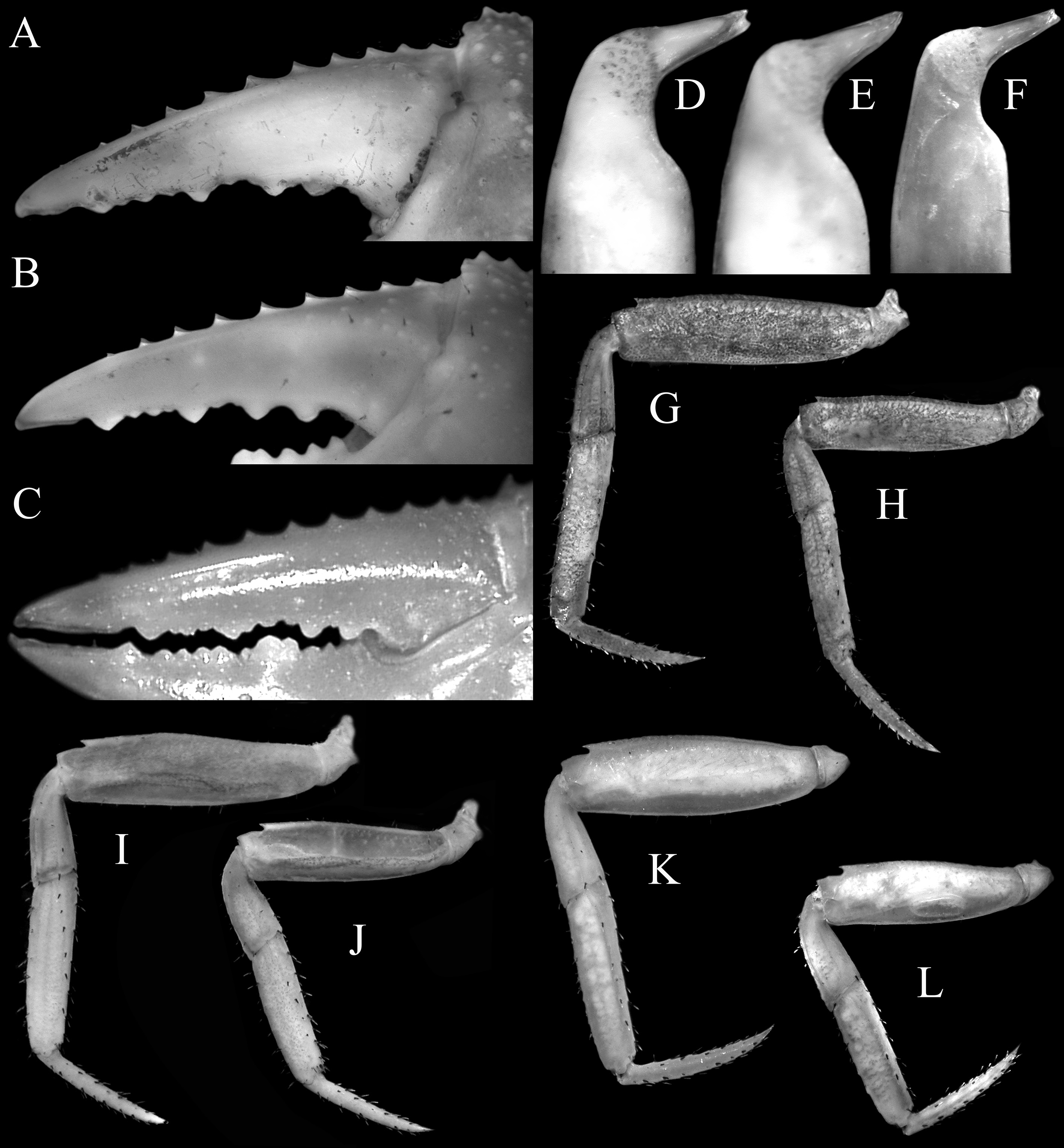
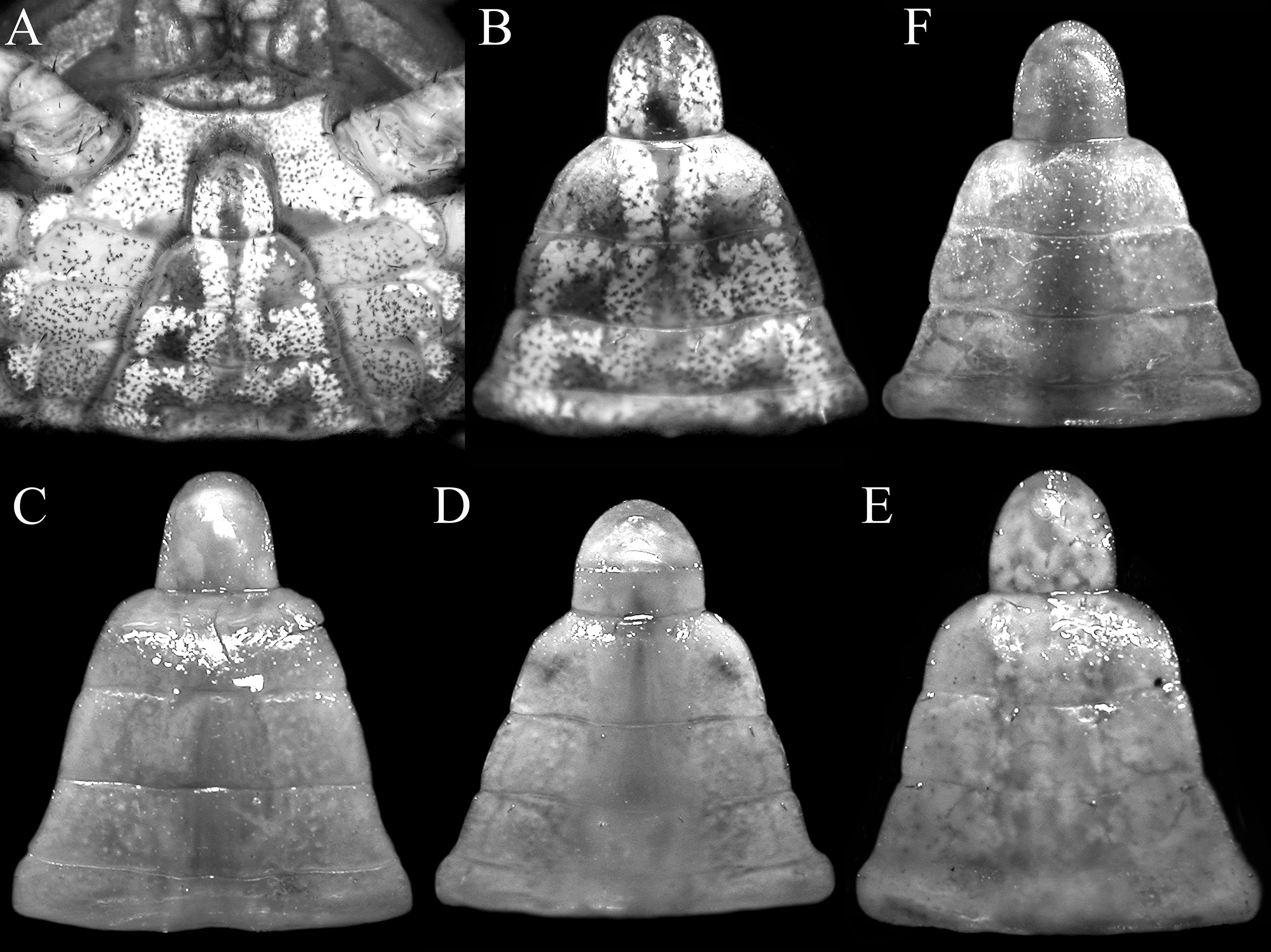
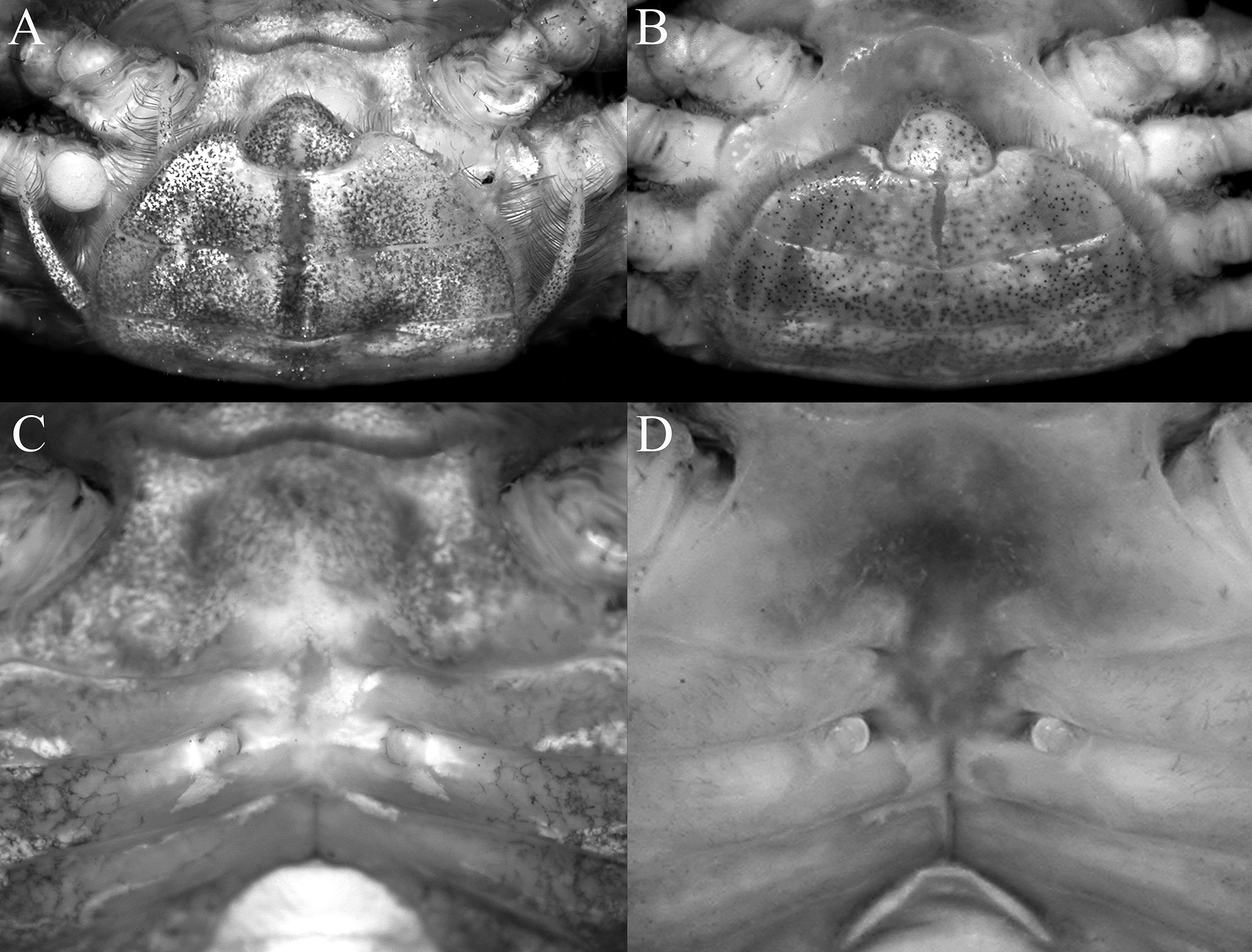
Diagnosis of male. Carapace quadrate, marginally wider than long or subequal, width to length ratio 1.01–1.06, lateral margins parallel, gently diverging towards posterior carapace margin ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 ); dorsal surfaces with well-defined regions, anterior half with numerous distinct tubercles of varying sizes, posterior half with smaller, lower tubercles ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 ); frontal margin distinctly deflexed, frontal lobes broad, with truncated margins in dorsal view, separated by shallow median concavity; postfrontal and postorbital cristae sharp, distinct ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 , 6A View FIGURE 6 ); external orbital angle triangular, directed obliquely anteriorly, clearly extending past lateral carapace margins, separated from first epibranchial tooth by distinct cleft; first epibranchial tooth low, blunt, second epibranchial tooth indistinct, separated from first by shallow notch ( Figs. 1A View FIGURE 1 , 5A View FIGURE 5 ); merus of third maxilliped rounded; exopod slender, flagellum absent ( Fig. 9A View FIGURE 9 ); outer surfaces of palm of chela covered with small rounded granules; dorsal margin of dactylus with 11 sharp, anteriorly directed tubercles ( Figs. 1C View FIGURE 1 , 6C View FIGURE 6 , 7A); ambulatory legs with long, slender merus with sharp subdistal spine on dorsal margin, surface weakly rugose, propodus slender, relatively long ( Figs. 1A View FIGURE 1 , 7G, H); pleon triangular; somite 6 broadest, with lateral margins gently convex; telson triangular, longer than broad, lateral margins gently convex ( Figs. 1B View FIGURE 1 , 8A, B); G1 stout, proximal, distal part bent at angle of ca. 45º from longitudinal axis ( Figs. 7D View FIGURE 7 , 9 View FIGURE 9 B–E), distal part elongate in lateral view, spatuliform from marginal view, with distinct median cleft on tip when viewed ventrally ( Figs. 7D View FIGURE 7 , 9D, E View FIGURE 9 ).
Females. Females resemble the males in all non-sexual characters, with chelae equal, relatively more slender, less inflated, tubercles on outer surface of palm and dorsal margin of dactylus smaller, less distinct ( Figs. 3A, B View FIGURE 3 , 5C View FIGURE 5 ). Pleon ovate, covering most of thoracic sternum ( Fig. 10A View FIGURE 10 ); vulvae positioned on anterior margin of thoracic sternite 6, with a round, stiff operculum, without sternal opercular cover ( Fig. 10C View FIGURE 10 ).
Variation. The G1s of a smaller paratype male (7.9 × 7.6 mm) agree well with that of the holotype. The external orbital angle is more slender, relatively shorter, blunter and does not extend past the lateral margins of the carapace, and the tubercles on the anterior half of the carapace are also less pronounced ( Fig. 5B View FIGURE 5 ). The chelae resemble those of the female detailed above.
Live colouration. In live specimens, the anterior half of the carapace is a deep, rusty, reddish-brown; the posterior half of the carapace and ambulatory meri being pale purple, while the carpi, propodi and dactyli are pale orange ( Figs. 1A View FIGURE 1 , 2A, 3A). The carpus, merus and palm of the chelipeds are bright purple in adult males, slightly duller in females and smaller specimens, with the fingers bright orange ( Figs. 1A, C View FIGURE 1 , 2B, 3B). The posterior half of the ambulatory legs are covered in white spots, which are faint to absent in large adult males but distinct in juveniles and all females ( Figs. 2C, D View FIGURE 2 , 4A, C View FIGURE 4 ). The white spots sometimes extend to the posterior half of the carapace in small specimens ( Figs. 2C, D View FIGURE 2 ). The ventral surfaces are a pale yellow with hints of purple, particularly on the telson ( Fig 1B View FIGURE 1 ). The eyes are olive green, and the eggs are orangish red ( Fig. 4B View FIGURE 4 ).
Etymology. From the Latin for a colorful image, alluding to the striking colour of the species in life. The name is used as a noun in apposition.
Remarks. In the structure of the G1, carapace features and colouration in life, Geosesarma spectrum n. sp. is closest to G. katibas Ng, 1995a , from central Sarawak. It can, however, be distinguished from the latter by the shape of the external orbital angle and epibranchial teeth, length of the ambulatory leg propodi, as well as the structure of the G1.
In G. spectrum n. sp., the external orbital angle is typically marginally narrower, and the cleft separating it from the first epibranchial tooth deeper ( Figs.1A View FIGURE 1 , 5 View FIGURE 5 A–C) (vs. marginally broader external orbital angle, shallower cleft separating external orbital angle from the first epibranchial tooth, Figs. 5D, E View FIGURE 5 ). The propodus of the ambulatory legs are longer ( Figs. 1A View FIGURE 1 , 7G, H) (vs. comparatively shorter in similar sized specimens, Figs. 7I, J View FIGURE 7 ). The proximal and distal parts of the G1 are less strongly bent when viewed laterally, and the tip of the distal part has a distinct median cleft, clearly visible from the ventral view ( Figs. 7D View FIGURE 7 , 9 View FIGURE 9 B–E) (vs. proximal and distal parts of G1 more strongly bent when viewed laterally, distinct median cleft at tip absent, Figs. 7E View FIGURE 7 , 9G, H View FIGURE 9 ). Geosesarma katibas was described from Ulu Katibas basin, Song District, Sarawak, with more recent material collected from Kanowit basin, Sarikei district, ca. 50 km away, two major downstream tributaries of the Rajang river. Given the limited distributional ranges of many Geosesarma , it can be assumed that the distribution does not overlap with G. spectrum n. sp., which have so far only been collected in several localities in Brunei, over 400km away.
Among the other known Bornean species of Geosesarma , the general structure of the G1, absence of flagellum on third maxilliped, and presence of a row of distinct tubercles on the dorsal margin of the cheliped fingers, ally Geosesarma spectrum n. sp. with G. bau Ng & Grinang, 2004 , G. gracillimum (De Man, 1902) and G. larsi Ng & Grinang, 2018 (from Sarawak) as well as G. ambawang Ng, 2015 , and G. pontianak Ng, 2015 (from Kalimantan Barat).
Geosesarma gracillimum View in CoL was originally described from Baram, Sarawak, and has been reported from other parts of northern Sarawak ( Holthuis 1979; Ng 2015). The species has previously been recorded from Brunei ( Choy 1991; Ng & Grinang 2004; Ng 2015). However, a more detailed examination of the Bruneian G. gracillimum View in CoL material shows that they all belong instead to G. spectrum View in CoL n. sp., thus restricting the distribution of G. gracillimum View in CoL to northern Sarawak.
Geosesarma spectrum View in CoL n. sp. differs from G. gracillimum View in CoL in having a relatively more narrow external orbital tooth that is directed obliquely anteriorly, with the outer margin distinctly convex, extending clearly past the lateral carapace margins ( Figs. 5 View FIGURE 5 A–C) (vs. a broader external orbital tooth that is directed obliquely outwards, with the outer margin straight to mildly convex, extending only slightly past lateral carapace margins, Fig. 5F View FIGURE 5 ); the distinctly more slender ambulatory meri ( Fig. 7G, H View FIGURE 7 ) (vs. proportionately broader ambulatory meri, Fig. 7K, L View FIGURE 7 ); and a slightly shorter and relatively stockier G1 with a distinct median cleft on the tip of distal part ( Figs. 7D View FIGURE 7 , 9 View FIGURE 9 B–E) (vs. proportionately longer, slenderer G1 with a shallower and less distinct median cleft on the tip of the distal part, Figs. 7F View FIGURE 7 , 9 View FIGURE 9 I–M).
Geosesarma spectrum View in CoL n. sp. can be distinguished from G. larsi View in CoL by possessing distinctly more numerous granules on the carapace surface ( Figs. 1A View FIGURE 1 , 5 View FIGURE 5 A–C) (vs. carapace surface with fewer granules, Ng & Grinang 2018: fig. 2A-C); the relatively shorter ambulatory meri and propodi ( Figs. 1A View FIGURE 1 , 7G, H) (vs. proportionately longer ambulatory meri and propodi, Ng & Grinang 2018: figs 2A, B, 3D–F); and a distinctly shorter and stouter G1 ( Figs. 7D View FIGURE 7 , 9 View FIGURE 9 B–E) (vs. relatively longer, more slender G1; Ng & Grinang 2018: fig. 5B–G).
Geosesarma spectrum View in CoL n. sp. differs from G. bau View in CoL in having the external orbital tooth longer and directed obliquely anteriorly ( Figs. 5 View FIGURE 5 A–C) (vs. external orbital tooth proportionately shorter and directed obliquely outwards, Ng & Grinang 2004: fig. 9A, B); relatively shorter ambulatory legs ( Figs. 1A View FIGURE 1 , 7G, H) (vs. proportionately longer ambulatory legs, Ng & Grinang 2004: figs. 8A, B); and a relatively stouter G1 with the distal part more strongly bent ( Figs. 7D View FIGURE 7 , 9 View FIGURE 9 B–E) (vs. more slender G1 with distal part less bent, Ng & Grinang 2004: figs. 9D–F).
Geosesarma spectrum View in CoL n. sp. can be separated from G. ambawang Ng, 2015 View in CoL , and G. pontianak Ng, 2015 View in CoL , in having the anterior carapace regions more granular ( Figs. 5 View FIGURE 5 A–C) (vs. anterior carapace region less granular, Ng 2015: figs. 8A, B, 11A, B); the merus of the third maxilliped is shorter than the ischium ( Fig. 9A View FIGURE 9 ) (vs. merus of third maxilliped equal or subequal to ischium, Ng 2015: figs. 9A, 13A); and the absence of a distinct subdistal hump on the G1 ( Fig. 9 View FIGURE 9 B–E) (vs. subdistal hump present on G1, Ng 2015: figs. 9D–I, 13D–H, J–M).
The live colour of adult males of G. spectrum n. sp. is almost identical to that of G. katibas ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 4A, B), and is diagnostic for these two species. Compared to G. spectrum n. sp., G. larsi is a deep purplish-red on the anterior half of the carapace, ambulatory meri and chelipeds ( Ng & Grinang 2018: fig. 1). Geosesarma ambawang and G. pontianak are a reddish-brown on the anterior half of the carapace and ambulatory legs, with the chelipeds a uniform bright red (Ng 2015: fig. 10). Geosesarma gracilimum is a uniform deep red to orange ( Fig. 4C, D View FIGURE 4 ) including the chelae, whereas G. bau have an orange to red carapace, with the chelae and legs brown (Ng 2015: 54, J. Grinang, pers. comm.).
Of the 13 species of Geosesarma now known from Borneo (including the present new one), only three have third maxilliped exopods which possess a long flagellum: G. amphinome (De Man, 1899) (Kapuas, West Kalimantan), G. pylaemenes Ng, 2015 (Anjungan, West Kalimantan), and G. sarawakense ( Serène, 1968) (western Sarawak) (see also Ng 2015: 51). The other 10 species which are distributed all over Borneo: G. ambawang Ng, 2015 , G. aurantium Ng, 1995b , G. bau Ng & Grinang, 2004 , G. danumense Ng, 2002 , G. gracillimum (De Man, 1902) , G. katibas Ng, 1995a , G. larsi Ng & Grinang, 2018 , G. pontianak Ng, 2015 , G. sabanum Ng, 1992 , and G. spectrum n. sp. have an exopod that completely lacks any trace of a flagellum. The reasons for this are not clear but may be associated with their habits. Those species with a well-developed flagellum on the exopod of the third maxillipeds may have more aquatic habits (it is certainly so for G. pylaemenes and G. sarawakense (see Ng 2015; unpublished data). From what is known (based mainly on available collection data), species which lack the flagellum do tend to be more terrestrial, climbing trees and can often be found far from streams ( Ng 1995a, b, 2015; Ebin & Chung 2012; Ng & Grinang 2018; Ng & Ng 2018; unpublished data). A detailed ecological study of all the species will need to be made to better understand these patterns and why the flagellum has been reduced or lost.
Ecological notes. The recent specimens of Geosesarma spectrum n. sp. were nocturnal, living in soil burrows on slopes, under well shaded canopy in primary rainforest within Ulu Temburong National Park and several hundred metres from any stream. Adult individuals were observed at night (ca. 8 pm) at the edge of their burrows, and more individuals, including juveniles, were found foraging on leaf litter and low foliage later in the night. A single juvenile was collected in pitcher plant, Nepenthes sp., suggesting juveniles may take refuge in phytotelmata instead of digging burrows. This is a frequently observed habit among many Geosesarma species (see Ng & Lim 1987; Ng 1988; Tan & Ng 2008). This has precedence, as one of the specimens (ZRC 2018.0746) was collected from a pitcher plant: “ Geosesarma is a wholly freshwater genus and some specimens, possibly of G. gracillima , were recently caught from the peat swamp at Seria (near the old railway). A few specimens were also collected from inside the pitcher of the pitcher plant Nepenthes bicalcurata [sic]. In this respect, G. gracillima from Seria is somewhat nepenthiphilous” ( Choy 1991: 144) (see also Clarke 1997).
One ovigerous adult female was observed wandering on leaf litter during the day. This specimen had its left eye damaged, and was probably attacked by a predator, and evicted from its burrow. The 15 eggs were eyed and believed to be close to hatching ( Fig. 4B View FIGURE 4 ). The number of eggs were probably more originally but some were probably lost during capture. The brooding female was maintained in captivity for 10 days, during which the eggs hatched, and several juvenile crabs were found clinging to the carapace of the mother ( Fig 4C, D View FIGURE 4 ). The juvenile crabs continued to cling to the mother for three days, leaving after completing their first molt. The present observation of the young crabs clinging to the carapace of the mother is notable, and known in several species, such as G. notophorum Ng & Tan, 1995 , from Pulau Lingga, Indonesia (cf. Ng & Tan 1995). This is the first confirmed record of this mode of brooding behavior for a Bornean species.







| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |
Geosesarma spectrum
| Ng, Paul Y. C. & Ng, Peter K. L. 2019 |
Geosesarma bau
| Ng, P. K. L. & Grinang, J. 2004: 1 |
Geosesarma cf. gracillima — Choy, 1991: 129
| Choy, S. C. 1991: 129 |