
Indalia predotae ( Schawerda, 1927 ) Macià & Ylla & Gastón & Huertas & Bau, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5191.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B66F9DFC-3BF3-42CA-B08F-F983FD615F4E |
|
DOI |
https://doi.org/10.5281/zenodo.7144145 |
|
persistent identifier |
https://treatment.plazi.org/id/039487AC-FFC2-BA68-FF11-E7A152F3FDB4 |
|
treatment provided by |
Plazi |
|
scientific name |
Indalia predotae ( Schawerda, 1927 ) |
| status |
comb. nov. |
Indalia predotae ( Schawerda, 1927) View in CoL comb. nov.
Original combination: Lithosia predotae Schawerda, 1927 . Zeitschrift des Österreichischen Entomologen-Vereines Wien 12: 37.
T. L.: Albarracín , (Aragón), Spain.
Material examined. SPAIN: HUESCA: 19 ♂ and 15 ♀, Tozal de la Mora, Villanueva de Sigena , 8.viii.1997, 3.viii.2002, 26.iv.2003, 31.v.2004, 10.ix.2005, 27.v.2006, 20.vi.2015, R. Macià & J. Ylla leg. ; 5 ♂ and 2 ♀, Barranco de Valcuerna , Candasnos, 295 m, 10.v.1993, 11.ix.2007, R. Macià & J. Ylla leg. ; 3 ♂, Ontiñena , 220 m, 30.viii.2008, R. Macià & J. Ylla leg. ; 4 ♂, Ontiñena , 215 m, 13.v.2015, J. Gastón leg. ; 2 ♂, Serreta Negra , Candasnos, 270 m, 3.x.2016, J. Gastón leg. ; TERUEL: 15 ♂ and 12 ♀, Camino rural de Moscardón a Royuela, Moscardón, 1446 m, 30TXK26, 4.ix.2013, R. Macià leg. ; 16 ♂ and 9 ♀ Valle de Valdevecar , Albarracín, 1125 m, 30TXK37, 15.v.1993, 27.viii.1995, 1.viii.1999, 3.ix.2013, 3.viii.2017, 11.viii.2018, R. Macià leg. ; 14 ♂ and 11 ♀, Camino Rural , Calomarde, 1550 m, 30TXK26, 2.viii.2017, R. Macià leg. ; ZARAGOZA: 6 ♂, Torralba de los Frailes, 1050 m, 27.vii.1999, J. Gastón leg. ; 15 ♂ and 8 ♀, Montes de la Retuerta, El Ciervo , 310 m, 30TYL39, 21.ix.2003, 24.ix.2005, 17.ix.2006, 19.ix.2014, R. Macià & J. Ylla leg. ; 18 ♂ and 14 ♀, Val de Boquero el Ciego, La Retuerta, 362 m, 30TYL29, 11.vi.2013, R. Macià leg.
Diagnosis. The grey-brownish wing tonality and the fringe between light yellow and ochre yellow make this species unmistakable. Indalia predotae is notably sexually dimorphic: the females are smaller, much lighter yellowbrown, and the wings are proportionally smaller in relation to the rest of the body.
Re-description. Imago ( Figs. 17–18 View FIGURES 17–24 ). Average wingspan males 23.4 mm (n=10; 20–27 mm); average wingspan females 19.1 mm (n=10; 17–21 mm).
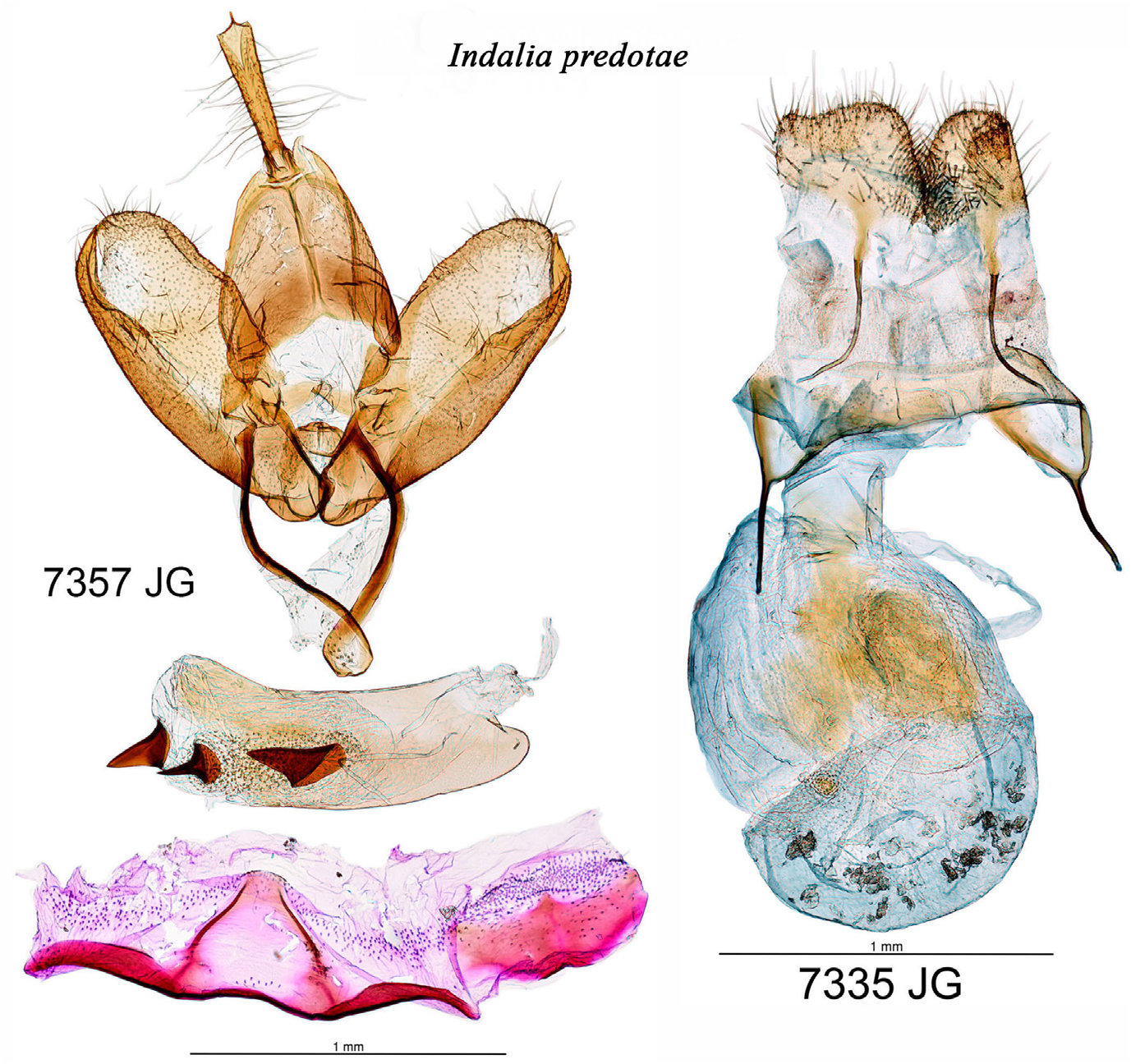
Genitalia ( Fig. 57 View FIGURE 57 ). Male genitalia: uncus cylindrical, slender, elongated, not flattened, sclerotised, with a pointed tip; valvae ovoid with rounded cucullus, sacculus wide and sclerotised, saccular process weakly sclerotised and slightly curved towards the cucullus. Aedeagus with three small, almost equal cornuti, two thick and coarse and the third smaller and sharper. Female genitalia: anal papillae small, posterior apophysis long, ostium and ductus bursae membranous; corpus bursa ovoid with one signum in its lower part and a weakly sclerotised plaque with diffuse edges in the place where the wart is located in other species. The appendix bursae prior to the ductus seminalis is thin and not very prominent.
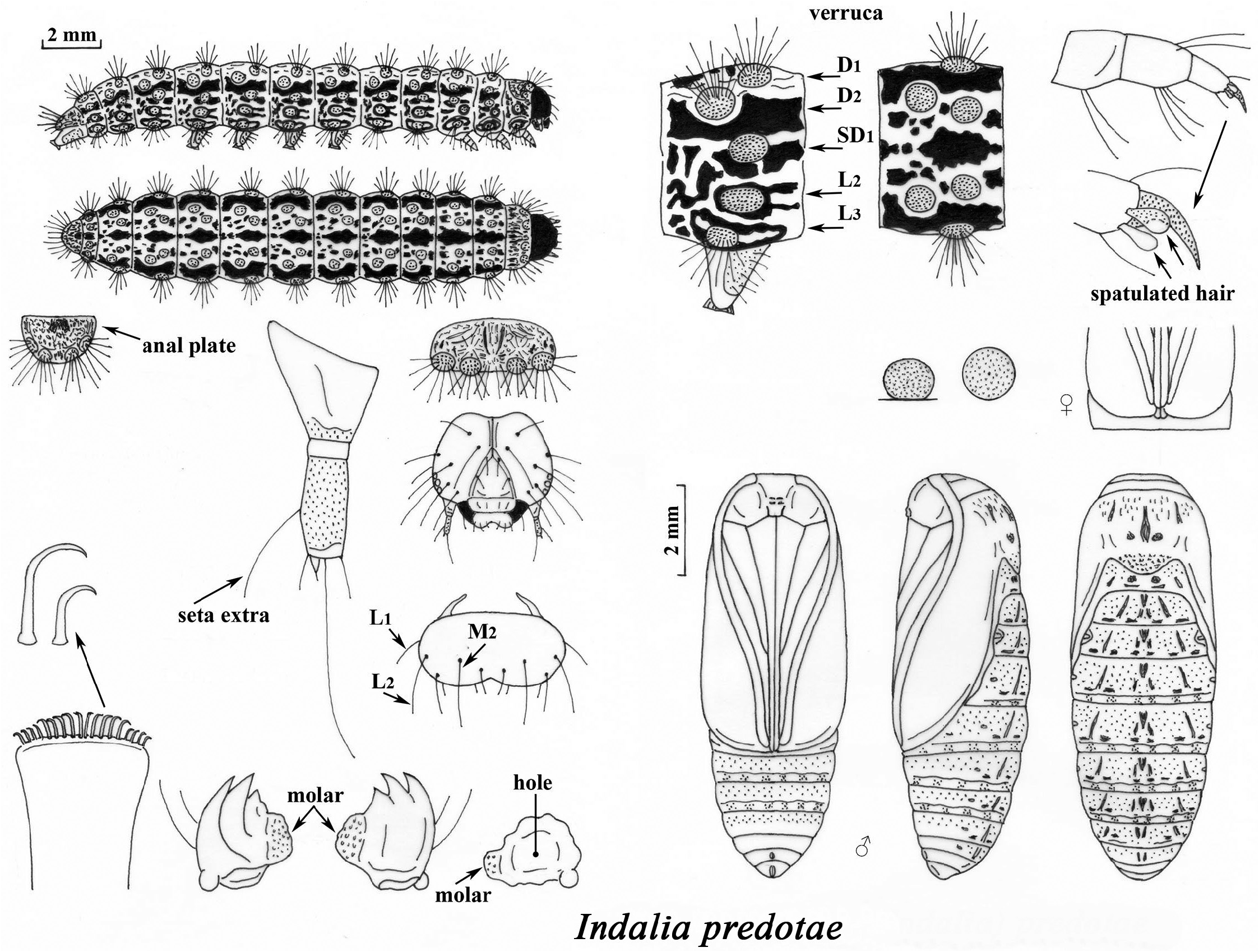
Immature stages ( Fig. 82 View FIGURE 82 ). The last instar larva is 17 mm long. Cephalic capsule completely black. Verrucae light brown (the L1 wart is missing or barely noticeable), all with translucent and black setae. Dorsal line broad at first and then narrow, broken, dark brown; broad light brown band in the area of D1 and D2 and then until SD 1 dark brown. Lateral sites to L3 light brown with various brown sinusoid spots. Pupa stylized, brown and smooth, with dark spots on the dorsal area; back of metanotum and segments 1 to 8, very small circular depressions; tip of abdomen rounded, without cremaster. In pupa of male, the antennae and proboscis reach the end of the wings, in the females they are shorter. The cocoon under lichens, bark and roks, with the silk very close to the body.
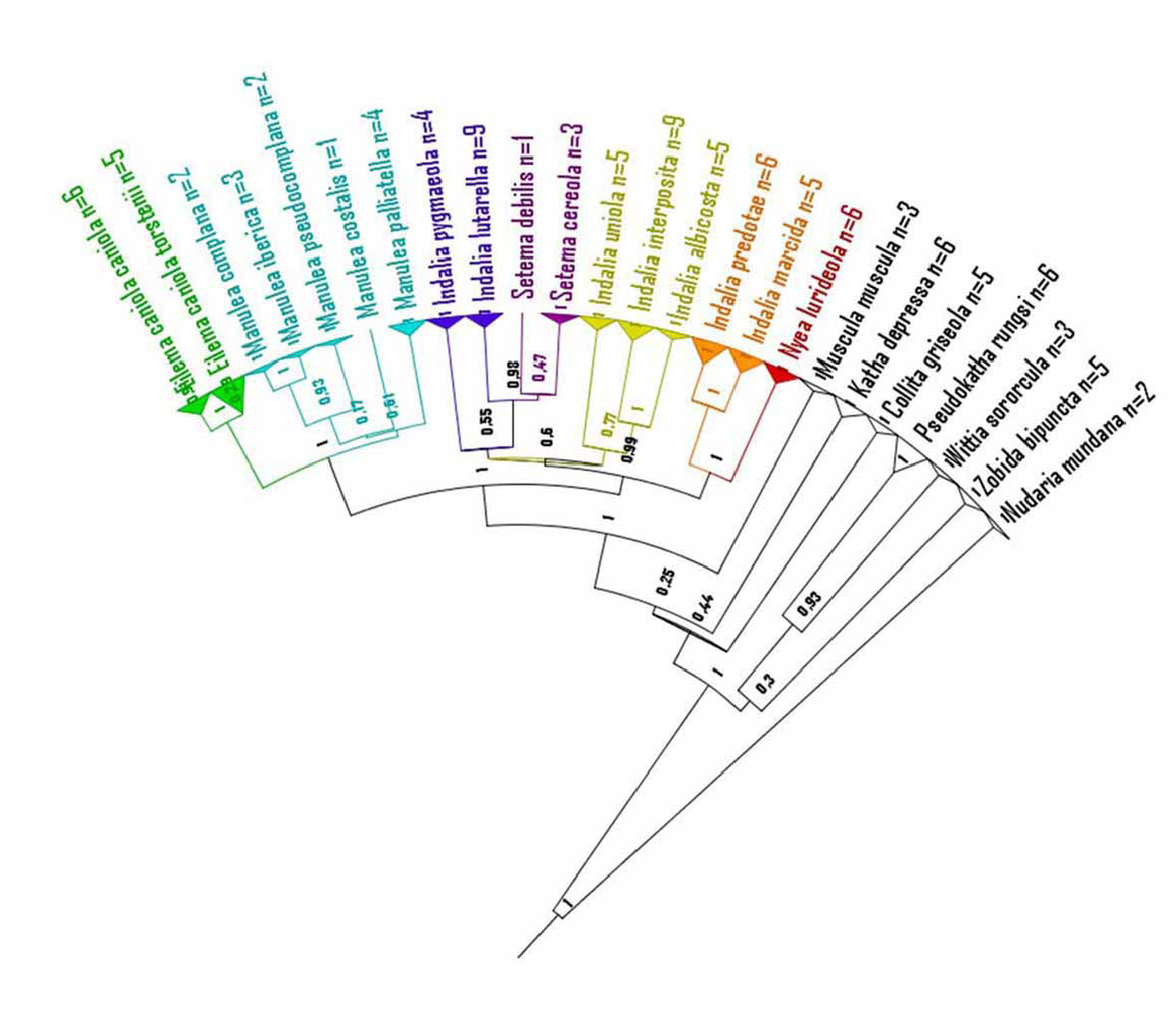
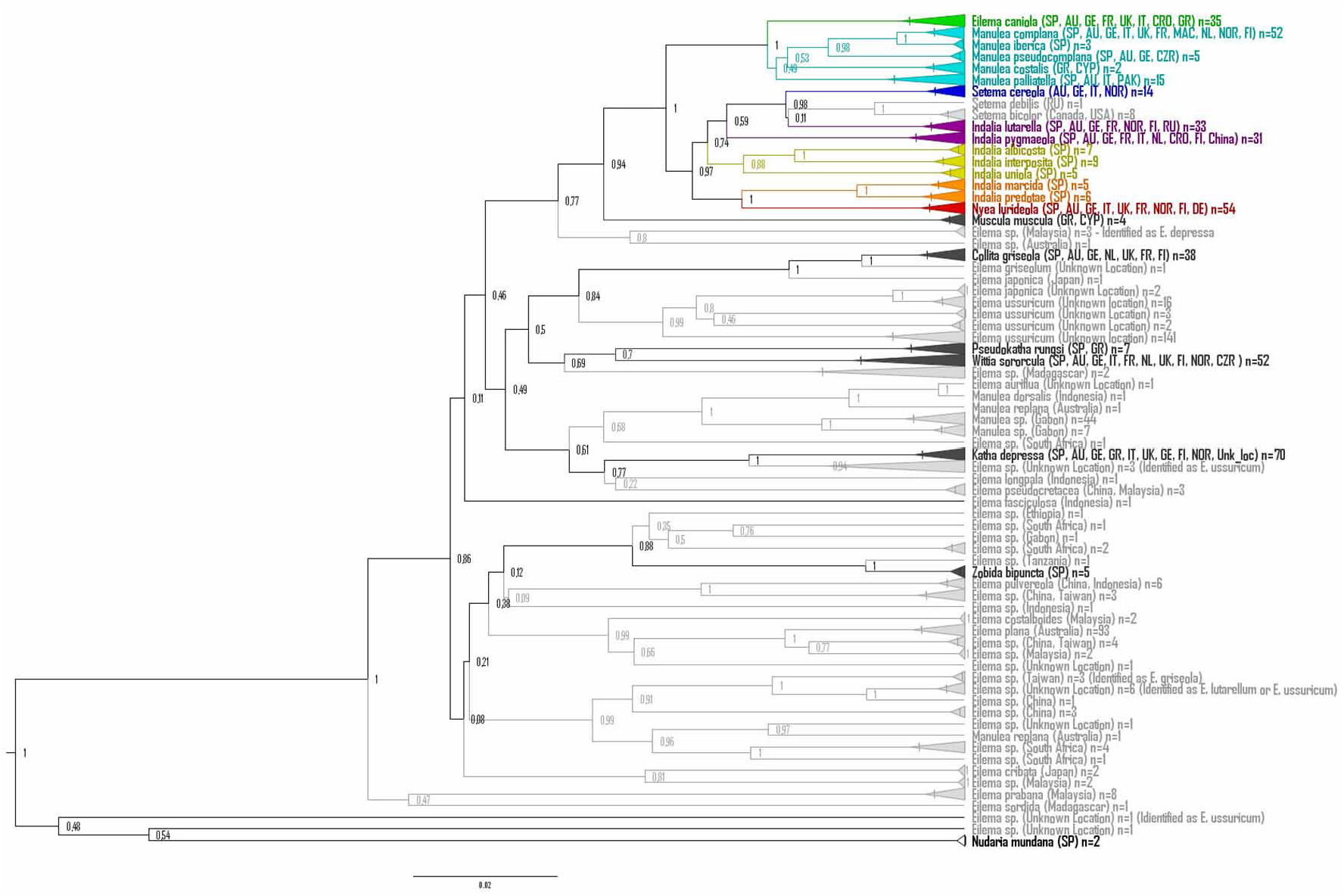
Molecular data. This species forms a clade with strong BI support (PP=1) and moderate within-group distance (0.47% ± 0.18SE). RESL Cluster Analysis returned a single cluster and all sequences coincided with a published BIN (BOLD: ACD0670). Like its close neighbour Indalia marcida, COI distances with other members of the genus are quite similar, albeit with lower values (TN-dist between 5.13% and 6.54%, average 5.42%) than those found for the latter (6.44% –7.97%; average 7.31%).
Biology. Bivoltine, with a first generation in May, June and July, followed by a second one in August, September and October. Xerothermic biotopes are favoured, with abundant steppe vegetation, and clearings in dry and sunny wooded areas among junipers and pine forests. Colonies are found up to 1200–1500 m above sea level. Adults are active both by day and at night, and are attracted by artificial light. The larvae feed on lichens, accepting dry lettuce leaves in captivity ( Ylla et al. 2010), and also prepared artificial diet.
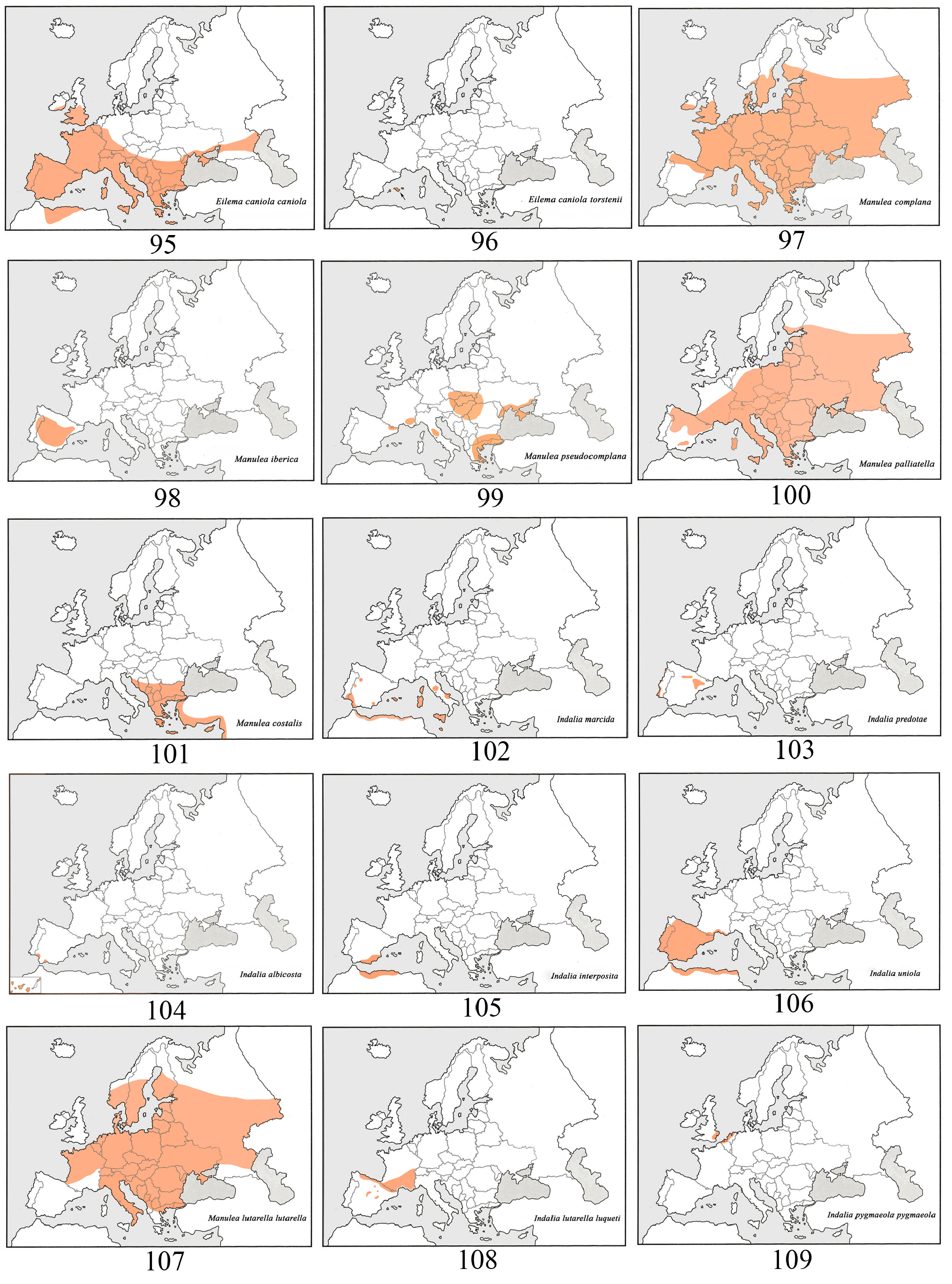
Distribution ( Fig. 103 View FIGURES 95–109 ). Iberian endemic. Known mostly from different localities in northeastern Spain, but absent from the entire Pyrenean area. The following are some examples of classic biotopes: Monegros and areas bordering the Ebro Valley, Iberian System (Albarracín and neighbouring areas) and La Vid (Burgos). Isolated colonies are known in Araba, Valladolid and Badajoz. Also known from southern and central Portugal (Teodoro Monteiro, 1975; Corley et al., 2000; Pires & Corley, 2007). Recently reported in Catalonia, Erms d´Aitona (Lleida), Guzmán (2015). Not known from the Balearic Islands.
The uniola group
Indalia uniola (Rambur, [1866]) comb. nov.
Indalia albicosta ( Rogenhofer, 1894) comb. nov.
Indalia interposita ( Rothschild, 1914) comb. nov.
It comprises three South European species: Indalia albicosta , endemic from the Canary Isles and also known from a few western Andalusian localities; Indalia uniola and Indalia interposita , which are spread along the North of Africa and reach southern Europe at different extent: Indalia uniola colonizes the Iberian Peninsula and south-east France, whereas Indalia interposita is restricted to the arid and desertic biotopes of south-east Spain. Genetically, the three species form a clade with moderately high Bayesian support (BI=0.77, Fig. 119 View FIGURE 119 ) which is confirmed and strengthened in the extended analysis (BI=0.88, Fig. 120 View FIGURE 120 ), and presents similarly low distances for both mitochondrial (aver. TN-dist=5.65%) and nuclear (2.08%) markers. Despite their genetic similarity, they show some phenotypic features that allow a fast and reliable morphological identification.

| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Indalia predotae ( Schawerda, 1927 )
| Macià, Ramon, Ylla, Josep, Gastón, Javier, Huertas, Manuel & Bau, Josep 2022 |
Lithosia predotae
| Schawerda 1927 |