
Copelatus cessaima, Caetano, Daniel S., Bená, Daniela De C. & Vanin, Sergio A., 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3710.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:FD7C6A48-0CA0-43F5-B937-59C3528F4769 |
|
DOI |
https://doi.org/10.5281/zenodo.6152286 |
|
persistent identifier |
https://treatment.plazi.org/id/038E87C0-FFE9-FFD2-FF3F-A1BDFBB5DECB |
|
treatment provided by |
Plazi |
|
scientific name |
Copelatus cessaima |
| status |
sp. nov. |
Copelatus cessaima View in CoL sp. nov.
(Figs. 1–8)
Type locality: Brasil, Parauapebas, Pará, Floresta Nacional de Carajás, 6°4'21.79"S 50°11'29.90"W.
Holotype: ♂, “ Brasil, Parauapebas, Pará, Floresta Nacional de Carajás, Gruta N4WS13 Flona, 6°4'21.79"S 50°11'29.90"W, XI/2006, Andrade et al. col.” Holotype label red, with black line border, printed: “ Holotypus Copelatus cessaima sp. nov., Caetano, Bená & Vanin det. 2013”. Deposited in MSZP.
Paratypes: Same data as holotype: 2 ♂ ( 1 ♂ dissected), 2 ♀ (1 ♀ dissected) (MZSP); same data as holotype: 2 ♂, 1 ♀ (MNRJ); same data, but 20.IV–04.V.2010: 1 ♂, 3 ♀ (MZSP); same data, but 10–19.V.2011: 1 ♀ (MZSP); same data, but Gruta N4E-22 Flona, 6°2'2.10"S 50°10'3.74"W, XI.2006: 1 ♂ dissected, 1 ♀ (MZSP); same data, but 7–12.X.2008: 1 ♂, 2 ♀ (MZSP); same data, but 20.IV–04.V.2008: 1 ♂ dissected (MZSP); same data, but 20.IV–04.V.2010: 3 ♀ (MZSP); same data, but Gruta N4E10 Flona, 6°2'20.16"S 50°9'38.28"W, XI.2006: 1 ♂ dissected, 1 ♀ (MZSP); same data, but 20.IV–04.V.2010: 3 ♀ (MZSP). All paratype labels red, with black line border, printed: “ Paratypus Copelatus cessaima sp. nov., Caetano, Bená & Vanin det. 2013”.
Measurements. Values are the min–max measurements from all type specimens followed by the measurement from the holotype specimen between parenthesis: total length 2.95–4.9 (3.9) mm; maximum width of the elytra 1.05–1.25 (1.25) mm; elytral length 2.05–2.4 (2.2) mm; maximum width of the pronotum 1.05–1.45 (1.2) mm; pronotum length 0.55–0.65 (0.59) mm.
Habitus. Head large, maximum width only slightly smaller than that of pronotum. Body flat, outline subparallel in dorsal view.
Color. Dorsal and ventral surface of integument usually light brown. Variation in color ranges from light brown to dark brown.
Surface sculpture. Dorsal surface without striae or strioles. Head with small punctures evenly distributed. Pronotum with a transversal punctate line of irregular dark maculae close to anterior margin; shape and number of maculae varying among specimens. Anterior quarter of elytron with dark punctate line parallel to internal margin of elytron, curved outwards near scutellum and ending at anterior margin of elytron.
Structural features. Eyes absent. Pronotum lacking lateral bead. Pronotum and elytra with long lateral setae. Pronotal setae restricted to anterolateral margin. Elytral setae regularly distributed alongside external margin in groups of two close setae (pairs); distance between pairs of setae twice as large as distance between setae within a pair (Fig. 1). Elytra completely fused. Metathoracic wings absent. Mentum and gula completely fused. Prosternal process lance-shaped, not protruded, not reaching metaventrite. Mesocoxae contiguous (Fig. 2). Anteromedial process of metaventrite short, apex rounded. Metacoxal lines strongly divergent anteriorly. Internal margin of anterior femur with row of setae, length of setae equal to half femur maximum width. External margin of anterior tibia with row of small spines, apical portion of internal margin with row of setae pointing forward, tip with three internal and three external spines. Internal margin of middle femur glabrous. External margin of mesotibia with row of spines, apical portion of internal margin with two spines, tip with two internal and two external spines. Segments I–III of pro- and mesotarsi ventrally with two pads formed by dense aggregation of setae in males (absent in females). Internal margin of hind femur glabrous. Hind tibia dorsoventrally flattened, tip with two internal and two external spines, internal dorsal-most spine longer than others (half-length of metatarsus I). Segments I–III of metatarsi with a fringe of long setae. Tarsal claws of all legs simple, slightly curved, with equal length. Notes: The large setae found on the lateral margins of the pronotum, elytra, and metatarsi (Fig. 1) were not found in some specimens, likely because these are very fragile structures that are easily broken during handling.
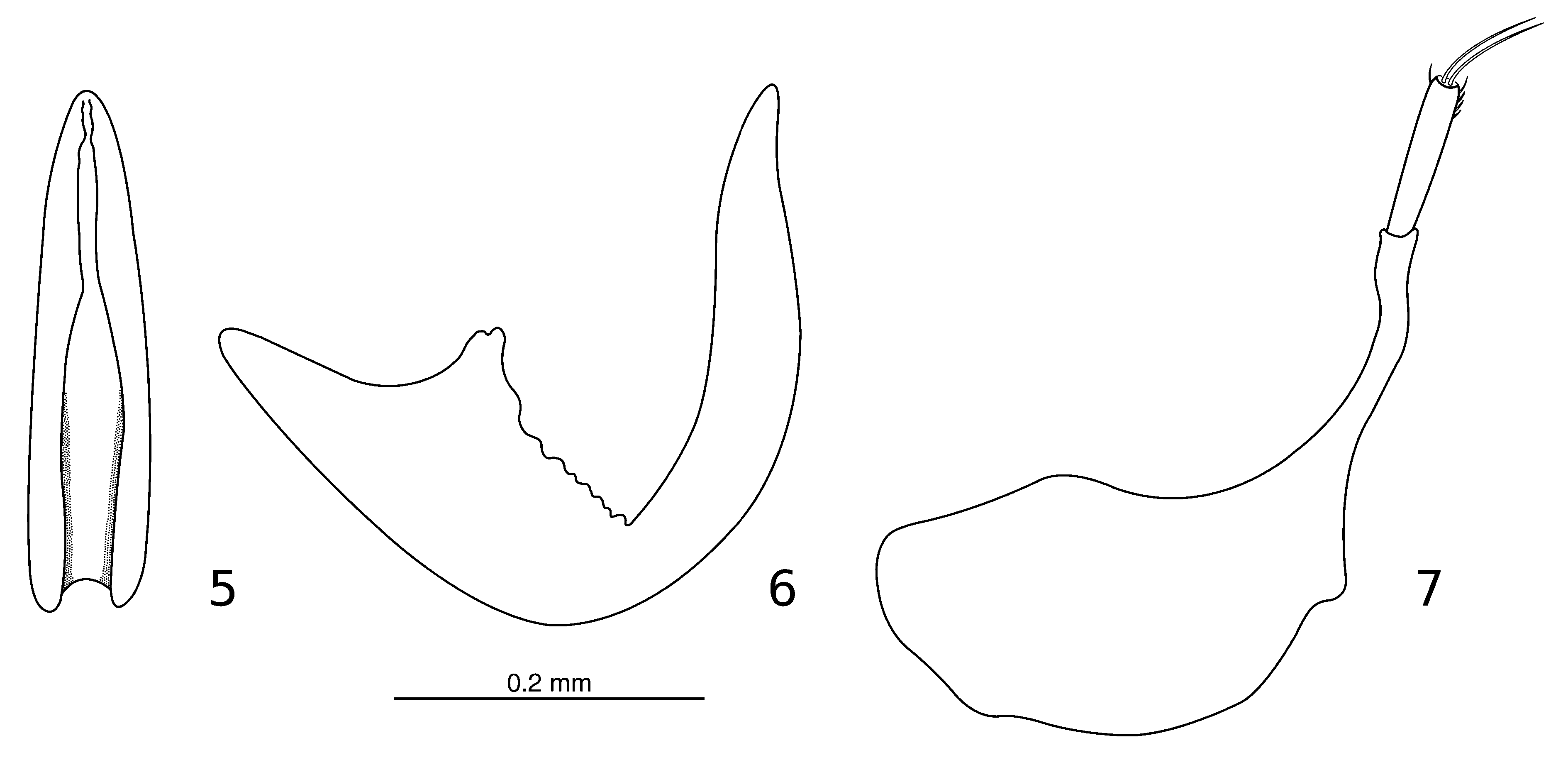
Male genitalia. Median lobe of aedeagus ( Figs. 5–6 View FIGURES 5 – 7 ) strongly curved, hook-shaped, becoming more slender towards anterior tip; basal portion of internal margin irregularly crenulated in lateral view ( Fig. 6 View FIGURES 5 – 7 ). Parameres ( Fig. 7 View FIGURES 5 – 7 ) symmetrical, broad at base, internal margin without setae; stylus straight, tip with row of four short setae on upper margin and a single short seta on lower margin, two sub-equal large apical setae.
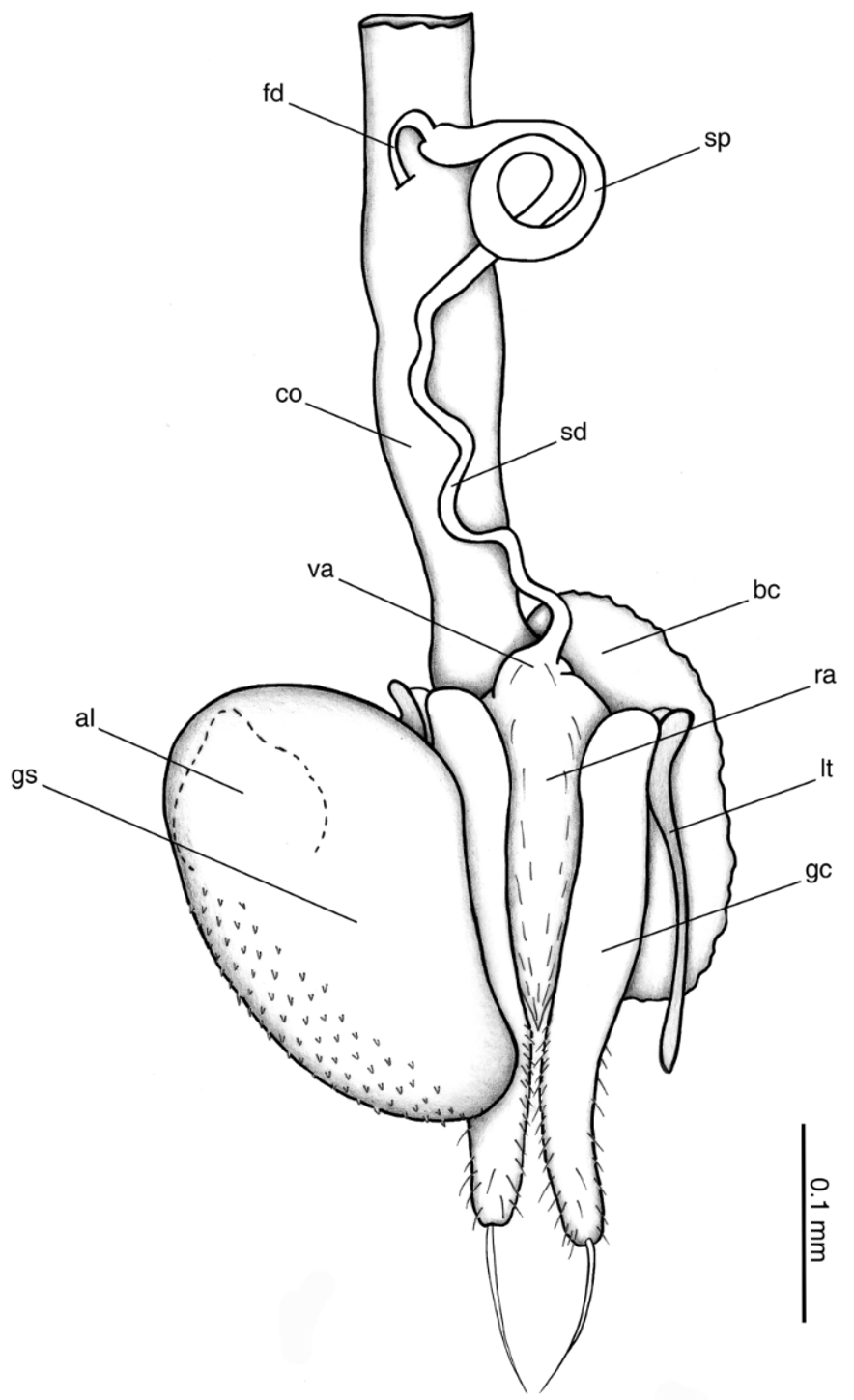
Female genitalia. Gonocoxa (gc) ( Fig. 8 View FIGURE 8 ) slender, lateral margins sinuous, broader near basal two thirds, distal third setose, apex with one long seta; laterotergite (lt) elongate, very slender; gonocoxosternite (gs) broadly oval, slightly narrowed towards apex, inner margin slightly concave, region near anterior outer margin with microtrichiae; anterior lobe of gonocoxosternite (al) nearly rounded with apex slightly protruded; bursa copulatrix (bc) present; spermatheca (sp) slender, loosely coiled, tapered to spermathecal duct (sd); fertilization duct (fd) broken, not observed (but see Fig. 8 View FIGURE 8 ).
Holotype features. The holotype does not differ from the paratypes in any of the described features. Etymology. The specific epithet “ cessaima ” is a non-latinized adjective, derived from the Tupi (Brazilian indigenous language) word "ceçaima", which means “blind” and refers to the eyeless condition of the species. Habitat. Specimens were found inside caves in water puddles naturally carved on iron rocks (ironstone formation).
Distribution. Only known from three caves of an ironstone formation at the Carajás National Forest, in the Brazilian municipality of Parauapebas, State of Pará, Brazil.
Troglomorphism and infrageneric systematic considerations
This troglomorphic species is readily differentiated from the other non-troglomorphic Copelatus species, for the eyes are absent, the mentum and the gula are fused, and the head is large. Exocelina abdita is the single other troglomorphic Copelatinae . Both species are similar in shape (Balke et al. 2004a, figs 2–3), but C. cessaima sp. nov. does not have the pronotal and elytral bases strongly narrowed, the lateral setae of the elytra are regularly distributed, and the metacoxal lines are divergent anteriorly.
Species restricted to caves and underground habitats commonly have reduced pigmentation and eyes reduced or completely absent. On the other hand, sensorial appendages are often highly developed, such as longer antennae and body setae, and longer and more slender legs (Romero 2004). Troglomorphic modifications in distantly related taxa can result in convergent and similar body shapes. These modifications, as pointed out by Balke et al. (2004a), can make it difficult to determine the correct taxonomic placement of a species. As a result, several monotypic genera defined due to their unique troglomorphic modifications have been synonymized with well-known genera. Copelatus cessaima sp. nov. is a species that lacks eyes, elytral striae, and metathoracic wings, and has completely fused elytra and long body setae (Figs. 1–4). These characters may lead to misplacement of C. cessaima sp. nov. in one of the 15 Copelatus species groups. In addition, these groups may be arbitrary subdivisions of the genus, given that they are not monophyletic (Miller 2001; Balke et al. 2004b). Therefore, here we did not assign C. cessaima sp. nov. to one of the Copelatus species groups, and assign the new species among those specified as “group unknown” by Nilsson (2013).
Acknowledgements
We thank R. Andrade for making the specimens available for study, R. Kawada for taking the specimen images and two anonymous reviewers for helpful comments. The equipment used to take the specimen photos was kindly provided by the Museu de Zoologia, Universidade de São Paulo. D.S. Caetano is supported by fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES—1093/12-6).
References
Balke, M., Watts, C.H.S., Cooper, S.J.B., Humphreys, W.F. & Vogler, A.P. ( 2004 a) A highly modified stygobiont diving beetle of the genus Copelatus (Coleoptera, Dytiscidae): taxonomy and cladistic analysis based on mitochondrial DNA sequences. Systematic Entomology, 29, 59–67.
http://dx.doi.org/10.1111/j.1365-3113.2004.00229.x
Balke, M., Ribera, I. & Vogler, A.P. ( 2004 b) MtDNA phylogeny and biogegraphy of Copelatinae, a highly diverse group of tropical diving beetles (Dytiscidae). Molecular Phylogenetics and Evolution, 32, 866 –880.
http://dx.doi.org/10.1016/j.ympev.2004.03.014
García, R., López, H. & Oromí, P. ( 2007) Additional data to the genus Baezia with description of a new species from a cave on El Hierro, Canary Islands (Coleoptera, Curculionidae, Molytinae). Zootaxa, 1631, 47–55.
Gnaspini, P., Vanin, S.A. & Godoy, N.M. ( 1998) A new genus of troglobitic carabid beetle from Brazil (Coleoptera, Carabidae, Zuphiini). Papéis Avulsos de Zoologia, 40 ( 19), 297–309.
Manivannan, D. & Madani, J.I. ( 2011) Two new species Hydrovatus sringeriensis and Copelatus wayanadensis (Coleoptera: Dytiscidae) from Western Ghats of South India. Records of the Zoological Survey of India, 111 ( 2), 1–6.
Miller, K.B. ( 1997) Reassignment of Agabus emines Kirsch to Aglymbus Sharp (Coleoptera: Dytiscidae) and redescription of the species. The Coleopterists Bulletin, 51 ( 3), 285–288.
Miller, K.B. ( 2001) On the phylogeny of the Dytiscidae (Insecta: Coleoptera) with emphasis on the morphology of the female reproductive system. Insect Systematics and Evolution, 32, 45–92.
http://dx.doi.org/10.1163/187631201x00245
Miller, K.B. & Balke, M. ( 2003) The unusual occurrence of aquatic beetles in amber, Copelatus aphroditae Balke, n. sp. and C. predaveterus Miller, n. sp., (Coleoptera: Dytiscidae: Copelatinae). Proceedings of the Entomological Society of Washington, 105 ( 4), 809–815.
Nilsson, A.N. ( 2007) Exocelina Broun, 1886, is the valid name of Papuadytes Balke, 1998. Latissimus, 23, 33–34.
Nilsson, A.N. ( 2013) A world catalogue of the family Dytiscidae, or the diving beetles (Coleoptera, Adephaga). University of Umeå, Sweden. Available from: http://www2.emg.umu.se/projects/biginst/andersn/index.htm (Accessed 11 March 2013)
Park, H.C., Lee, Y.B. & Lee, H.A. ( 2008) Taxonomic review of the genus Copelatus Erichson (Coleoptera: Dytiscidae) in Korea. Entomological Research, 38, 73–76.
http://dx.doi.org/10.1111/j.1748-5967.2008.00134.x
Romero, A. ( 2004) Evolution of hypogean fauna. In: Gunn, J. (Ed.), Encyclopedia of Cave and Karst Science. New York: Fitzroy Dearborn, pp. 347–349.
Yin, Z.W., Li, L.Z. & Zhao, M.J. ( 2011) Discovery in the caves of Guangxi, China: three new troglobitic species of Tribasodites Jeannel (Coleoptera, Staphylinidae, Pselaphinae). Zootaxa, 3065, 49–59.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Copelatinae |
|
Genus |